
A veszélyeztetett, magas THC-tartalmú kannabisz (Cannabis sativa subsp. indica) domesztikátumainak és vad rokonainak osztályozása
Eredeti angol cím: A classification of endangered high-THC cannabis (Cannabis sativa subsp. indica) domesticates and their wild relatives
Szerzők: John M. McPartland‡§, Ernest Small|
Intézményi hovatartozások: ‡ GW Pharmaceuticals, Cambridge, Egyesült Királyság; § University of Vermont, Burlington, Amerikai Egyesült Államok; | Agriculture and Agri-Food Canada, Ottawa, Kanada
Levelező szerző: John M. McPartland (mcpruitt@myfairpoint.net)
Tudományos szerkesztő: Hugo de Boer
Előzménydátumok: beérkezett: 2019. szeptember 19.; elfogadva: 2020. február 2.; megjelent: 2020. április 3.
McPartland & Small: a high-THC kannabiszformák botanikai osztályozása
Ez a tanulmány a háromrészes rendszertani sorozat botanikai kiindulópontja. McPartland és Small nem a mai kereskedelmi „Sativa” és „Indica” címkéket próbálja igazolni, hanem azt vizsgálja, milyen történeti, földrajzi, morfológiai és fitokémiai alapon különíthetők el a nagy THC-tartalmú kannabisz régi dél-ázsiai és közép-ázsiai alakkörei, valamint azok vadon növő rokonai.
A szerzők a köznyelvben „Sativa”-ként emlegetett dél-ázsiai drogtípusú domesticátumot Cannabis sativa subsp. indica var. indica néven, a közép-ázsiai, afgán jellegű domesticátumot pedig Cannabis sativa subsp. indica var. afghanica néven értelmezik. Ezek mellé két vad jellegű rokonformát is társítanak: a dél-ázsiai/himalájai var. himalayensis és a közép-ázsiai var. asperrima taxont. A cikk így nem egyszerűen két piaci kategóriáról, hanem négy, megőrzési szempontból is fontos botanikai-génanyagbeli egységről beszél.
A tanulmány jelentősége abban áll, hogy a kannabisz régi rendszertani neveit és a modern piaci szóhasználatot szétválasztja egymástól. A mai „Sativa”, „Indica” és „Hybrid” jelölések sokszor hibrid, dokumentálatlan nemesítési hátterű növényeket takarnak, míg a tanulmányban tárgyalt taxonok történeti tájfajtákhoz, földrajzi eredethez, morfológiai különbségekhez és fitokémiai mintázatokhoz kötődnek.
Hivatkozás: McPartland JM, Small E (2020) A classification of endangered high-THC cannabis (Cannabis sativa subsp. indica) domesticates and their wild relatives. PhytoKeys 144: 81–112. DOI: 10.3897/phytokeys.144.46700
Eredeti cikk: https://phytokeys.pensoft.net/article/46700/
Szerzői jog és licenc: © 2020 John M. McPartland, Ernest Small. Ez nyílt hozzáférésű cikk, amely a Creative Commons Attribution License (CC BY 4.0) feltételei szerint terjeszthető; ez korlátlan felhasználást, terjesztést és bármely médiumban történő reprodukciót engedélyez, feltéve, hogy az eredeti szerzőt és forrást feltüntetik.
Magyar fordítás és szerkesztési megjegyzés: Magyar fordítás és ábraszöveg-adaptáció. A fordítás az eredeti szerzők, a forrás, a DOI és a licenc feltüntetésével készült; a változtatás a magyar nyelvű fordítás és szerkesztés.
Kapcsolódó tanulmányok a sorozatban: Genomikai-taxonómiai áttekintés 2023 · Pángenomikai-evolúciós értelmezési modell 2025
Tartalomjegyzék
Kivonat
A drog-típusú Cannabis két fajtája az 1980-as években kapott laikus elnevezéseket. A „Sativa” Dél-Ázsiából (Indiából) származott, korai történeti elterjedéssel Délkelet-Ázsiába, Afrikába és Amerikába. Az „Indica” Közép-Ázsiából (Afganisztánból, Pakisztánból, Turkesztánból) eredt. E változatoknak egyértelmű taxonómiai neveket adtunk, miután 1100 herbáriumi példány morfológiai bélyegeit megvizsgáltuk, valamint a szakirodalomból származó fitokémiai és genetikai adatokat meta-analízisben elemeztük. A „Sativa” és az „Indica” rendre C. sativa subsp. indica var. indica és C. sativa subsp. indica var. afghanica néven kerül elismerésre. Vadon növő rokonaik a C. sativa subsp. indica var. himalayensis (Dél-Ázsiában) és a C. sativa subsp. indica var. asperrima (Közép-Ázsiában). A divergenciát a természetes szelekció indította el, amelyet Dél- és Közép-Ázsia éghajlati viszonyai hajtottak. A későbbi domesztikáció további fitokémiai divergenciát eredményezett. A dél- és közép-ázsiai domesztikátumok megkülönböztethetők tetrahidrokannabinol- és kannabidiol-tartalmuk (THC/CBD arányok, rendre ≥7 vagy <7), terpenoidprofiljuk (a szeszkviterpén-alkoholok hiánya vagy jelenléte), valamint morfológiai bélyegeik együttese alapján. A két domesztikátum az elmúlt 50 évben kiterjedt introgresszív hibridizáción ment keresztül. Ez eltüntette a jelenleg hozzáférhető, hibridizált „Sativa” és „Indica” közötti különbségeket. Azok a „fajtavonalak” („strains”), amelyek állítólag a „Sativa” és az „Indica” képviselői, rendszerint dokumentálatlan hibrid hátterű növények THC/CBD arányain alapulnak (az úgynevezett „Indica”-kat gyakran egyszerűen az alapján határolják le, hogy több CBD-t tartalmaznak, mint a „Sativa”-k). Az itt bemutatott osztályozás a Cannabis négy olyan taxonját határolja körül és nevezi meg, amelyek a modern kannabinoid fajtavonalak eredetét adó, kritikusan veszélyeztetett génanyag-rezervoárokat képviselnek, és sürgős megőrzésre szorulnak.
Kulcsszavak
Kannabinoidok, Cannabis sativa, osztályozás, ökológia, génanyag, marihuána, nevezéktan
Bevezetés
A Cannabis ősi domesztikátum, háromféle hasznosítású kultúrnövény. A régészek táplálkozási kontextusban, egy konyhai hulladékhalomban találtak terméseket („magokat”), 8000 kalibrált évvel i. e. keltezett radiokarbon dátummal (Kudo et al. 2009). A rosthasználat bizonyítéka majdnem ugyanilyen régi, bár az ősi zsinórozat Cannabis-ként való azonosítása (vagy ugyanennek kerámialenyomatai alapján) némileg szubjektív (McPartland and Hegman 2018). Drogkontextusból származó leletek – kannabinoidokat tartalmazó, megégett maradványok egy füstölőedényben – i. e. 500 kalibrált évre keltezhetők (Ren et al. 2019). A Cannabis korai megnevezései közé tartozik a kínai má, amely kb. i. e. 750–600 között adatolt (Qu and Waley 1955), a qunubu, egy újasszír jövevényszó a szkíta nyelvből, kb. i. e. 680-ból (Seidel 1989), valamint a κάνναβις, egy görög jövevényszó a szkítából, kb. i. e. 440-ből (Herotodus 2007).
A latin Cannabis sativa nevet rendszerint Leonhart Fuchsnak tulajdonítják, a binomiális nevet azonban valójában Ermolao Barbaro alkotta meg 1480 és 1490 között; műve 23 évvel a halála után jelent meg (Barbaro 1516). Carl Linné átvette a binomiális nevet a Species Plantarum-ban, amely a botanikai nevezéktan nemzetközileg elismert kezdőpontja (Linnaeus 1753). Jean-Baptiste Lamarck szakított a linnéi ortodoxiával, amikor a drog-típusú növények számára egy második fajt, a C. indica-t ismerte el (Lamarck 1785).
Small and Cronquist (1976) egyfajos koncepciót javasoltak. Linnaeus és Lamarck taxonjait alfaji rangon választották szét C. sativa subsp. sativa és C. sativa subsp. indica (Lam.) E. Small & Cronq. néven. Az alfajokat a ∆9-tetrahidrokannabinol (THC) tartalom alapján határolták körül. A C. sativa subsp. sativa-t úgy definiálták, mint amely a nőivarú (pisztillátus) növények szárított virágzó csúcsaiban <0,3% THC-t tartalmaz, a C. sativa subsp. indica-t pedig úgy, mint amely ≥0,3% THC-t tartalmaz. Számos ország beépítette a 0,3%-os kritériumot a rosttípusú (kender) növényeket és a drog-típusú (marihuána) növényeket szabályozó előírásokba.
Egyes botanikusok előnyben részesítik a C. sativa L. és a C. indica Lam. faji rangon történő elismerését (Hillig 2005a, Clarke and Merlin 2013). A taxonómiai rangról szóló viták közismerten önkényesek. A DNS-szekvenciákat alkalmazó molekuláris vizsgálatok kevésbé önkényessé tehetik a rang kérdését. Mandolino et al. (2002) tíz drog- és rosttípusú változatban számszerűsítették a DNS-polimorfizmusokat. Több variabilitást találtak egy adott változaton belüli egyedek között, mint a változatok között – olyan adatokat, amelyek megerősítették „egy egységes, széles körben közös génkészlet” létét. A Cannabis világméretű gyűjteményében Gilmore et al. (2007) alacsony szekvenciavariációs rátát találtak (körülbelül 1 polimorfizmus 1 kb szekvenált cpDNS-re) – ami egyetlen fajjal konzisztens.
McPartland (2018) DNS-vonalkódokat használt mérőszámként arra, hogy a Cannabis rangjának kérdését más növények kontextusába helyezze. Öt növényi vonalkódot vizsgált (rbcL, matK, trnH-psbA, trnL-trnF és ITS1), és 0,41%-os átlagos divergenciát (vonalkód-rést) számított a C. sativa és a C. indica között. Ez majdnem megegyezett öt olyan növénypár 0,43%-os átlagos divergenciájával, amelyeket különböző változatoknak vagy alfajoknak tekintenek (pl. Camellia sinensis var. sinensis és C. sinensis var. assamica). Ezzel szemben 3,0%-os vonalkód-rés választott el öt olyan növénypárt, amelyeket külön fajoknak tartanak (pl. Humulus lupulus és H. japonicus). Hebert et al. (2004) a két COI-szekvencia közötti 2,7%-os különbséget (a „vonalkód-rést”) javasolták küszöbértékként arra, hogy a genetikailag divergáló példányokat külön állatfajként jelöljék meg.
Sawler et al. (2015) 0,156-os átlagos fixációs indexet (FST) számítottak rost- és drog-típusú növények populációi között (n = 43, illetve 81). Az FST értékek 0 és 1 között mozognak; a nulla érték azt jelzi, hogy a két csoport szabadon kereszteződik, az 1-es érték pedig azt, hogy a csoportok egymástól teljesen izoláltak. A 0,156-os átlagos FST hasonló ahhoz a genetikai differenciálódási fokhoz, amely az Európában és Kelet-Ázsiában élő emberi populációk között van; ezek egyetlen fajhoz tartoznak.
Lynch et al. (2016) FST = 0,099 értéket számítottak a rost- és drog-típusú csoportok között (n = 22, illetve 173). Grassa et al. (2018) FST = 0,229 értéket számítottak a rosttípusú hozzáférések és a „marihuána” között, Sawler, Lynch és saját szekvenálási adataik összefűzésével. Hey and Pinho (2012) az FST = 0,35 értéket javasolták konzervatív küszöbmérőszámként a faji differenciálódáshoz; az ennél nagyobb értékű párokat külön fajokként, a kisebb értékű párokat alfaji populációkként azonosítják. Nyilvánvaló, hogy a C. sativa L. és a C. indica Lam. legjobban alfaji rangon különíthető el.
Az 1980-as években a drog-típusú növényeket két kategóriára kezdték osztani, amelyeket mindenütt elterjedt címkékkel „Indica” és „Sativa” néven ismertek. Ez a népi taxonómia azután vált széles körben elterjedtté, hogy Anderson (1980) közzétette a növények vonalas rajzát (1. ábra). Az „Indica” és a „Sativa” megkülönböztetését morfológia és földrajzi eredet alapján végezte. Ahogyan de Meijer and van Soest (1992) összefoglalták, az „Indica” széles levélkéjű, alacsony és kompakt habitusú, korai érésű növényekre vonatkozott, és van bizonyíték arra, hogy az ilyen növények tájfajta ősei Közép-Ázsiából (elsősorban Afganisztánból) származtak. A „Sativa” keskeny levélkéjű, magas és laza habitusú, késői érésű növényekre vonatkozott, és van bizonyíték arra, hogy az ilyen növények tájfajta ősei eredetileg Dél-Ázsiából (elsősorban Indiából) származtak, korai történeti elterjedéssel Délkelet-Ázsiába, Afrikába és Amerikába.
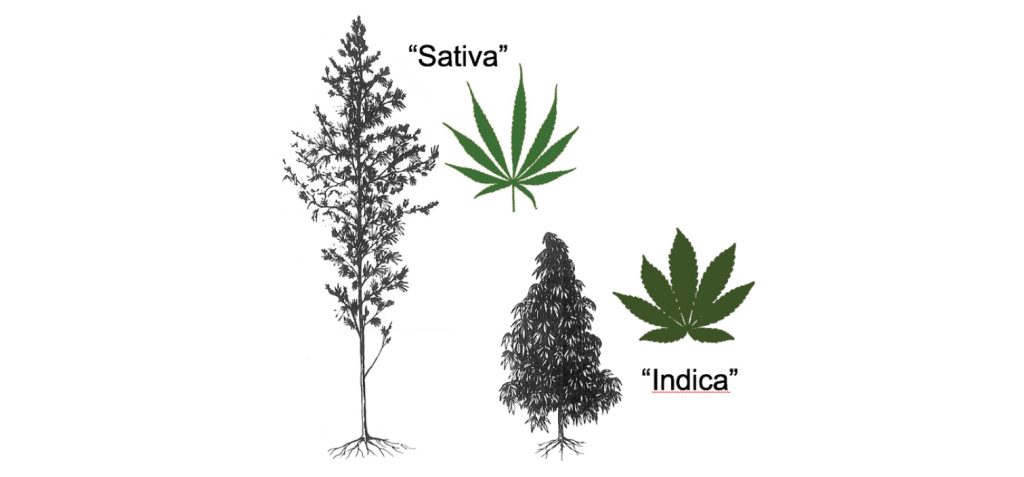
Clarke (1981) elfogadta Anderson „Indica” koncepcióját a Közép-Ázsiából származó növényekre: „Az e területről származó fajtavonalakat gyakran használják a Cannabis indica típuspéldáiként.” A morfológiai különbségeken túl egy fitokémiai bélyeget is megjegyzett – a közép-ázsiai növények egyedülállóan csípős, „skunk”-szerű aromát termeltek. Clarke (1987) egy organoleptikus minőséget is hozzátett – az Afganisztánból származó növények „lassú, lapos, nyomasztó betépést” idéztek elő. Hillig (2005a) a közép-ázsiai tájfajtákat széles levélkeátmérőjű (WLD) biotípusokként, a dél-ázsiai örökségű tájfajtákat pedig keskeny levélkeátmérőjű (NLD) biotípusokként említette. A WLD és NLD biotípusok genetikában (Hillig (2005a)), morfológiában (Hillig 2005b), THC-kannabidiol (CBD) arányokban (Hillig and Mahlberg 2004) és terpenoidtartalomban (Hillig 2004) különböztek.
Újabb szerzők tévesen azonosították a népi „Sativa” kifejezést a C. sativa tudományos név epithetonjával, és tévesen azonosították a népi „Indica” kifejezést a C. indica tudományos név epithetonjával; ezeket az eltéréseket először McPartland et al. (2000) jegyezték meg. Small (2007) kijelentette, hogy a „Sativa” és az „Indica” „meglehetősen inkonzisztens” a formális nevezéktannal. Linnaeus C. sativa típuspéldánya rosttípusú (kender) növény, nem drog-típusú (marihuána), ezért a „Sativa” kifejezést helytelenül alkalmazták drog-típusú növényekre (logikusan a rosttípusú kender számára kellene fenntartani). Lamarck a C. indica-t Indiából származó drog-típusú növényekre, valamint délkelet-ázsiai és afrikai leszármazottaikra írta le – ezeket ma ellentmondásos módon „Sativa”-nak nevezik (logikusan az „Indica” kifejezést a Lamarck által leírt drognövények számára kellene fenntartani).
A népi „Sativa” (amely a többnyire vagy teljesen THC-t tartalmazó kannabinoidprofilú növényeket jelöli) és a „C. sativa” (szűk nevezéktani értelemben az alacsony THC-tartalmú kenderformákat jelölve), továbbá a népi „Indica” (amely jelentős THC-t, de gyakran jelentős CBD-t is tartalmazó növényeket jelöl) és a „C. indica” (szűk nevezéktani értelemben magas THC-, alacsony CBD-tartalmú formákat jelölve) téves megfeleltetései taxonómiai tanulmányokban és jogi dokumentumokban is megjelentek. Még a „Nature” oldalait is problematikusan díszítette a „Sativa” és az „Indica”, az 1. ábra egy változatával együtt (Gould 2015). A biológiai osztályozás összetettségét és finomságait nem ismerőket ez félrevezetheti, de az alapelv egyszerű: a „Sativa” és az „Indica” kifejezéseket kétértelműen és ellentmondásosan használták.
A múlt évszázadokban a dél-ázsiai örökségű tájfajtákat világszerte sokkal szélesebb földrajzi tartományban termesztették, mint a közép-ázsiai tájfajtákat. Utóbbiak csak az 1970-es évek elején kerültek a nyugati Cannabis-nemesítők figyelmébe. Azóta a nemesítők esetlegesen hibridizálták a közép-ázsiai és dél-ázsiai tájfajtákat, és nagyrészt eltüntették fenotípusos különbségeiket (Clarke and Merlin 2013; Small 2017). Már 35 évvel ezelőtt is nehézzé vált hibridizálatlan tájfajtákhoz jutni az USA-ban és Európában (Clarke 1987). A „Sativa” és „Indica” hibridek rendkívül népszerűnek bizonyultak. Az „Indica” gének hasznosak a kannabinoidhozam növelésére, a magas földrajzi szélességeken szabadföldi növények érésének gyorsítására, valamint a növénymagasság csökkentésére, hogy azok könnyebben elrejthetők legyenek kültéren és könnyebben termeszthetők legyenek beltéren. A gyorsan növekvő CBD-piacon az „Indica” gének (gyakran tévesen „Ruderalis”-ként címkézett növényekből) növelték a CBD arányát a THC-hoz képest a növényi termékekben.
Riasztó módon a közép- és dél-ázsiai tájfajtákat idegen génanyag bevitele rontotta meg saját diverzitási központjaikban. Beisler (2006) azzal dicsekedett, hogy 1972 körül „Mexican Gold”-ot vitt Afganisztánba. Casano (2005) megjegyezte, hogy az afgán tájfajták más drog-típusú növényekkel való hibridizáció miatt „eltűnőben” voltak. Fordítva, közép-ázsiai tájfajtákat vezettek be a dél-ázsiai diverzitási központokba az 1970-es években – Nepálba (Cherniak 1982), Jamaicába (Lamb 2010) és Thaiföldre (Clarke and Merlin 2016). 1980-ra afgán tájfajtákat importáltak Dél-Kasmírba, rostált hashīsh előállítására termesztették, és a termesztésből kiszökött egyedek a szántóföldek közelében nőttek (Clarke 1998). Ugyancsak az 1980-as években közép-ázsiai genetikát vittek be Dél-Afrikába (Peterson 2009) és Marokkóba (Clarke and Merlin 2016). Sharma (1988) a Himachal Pradesh állambeli Kulluban növő „hibrid Cannabis”-ról írt, és „külföldi állampolgárokat” tett felelőssé.
A közép- és dél-ázsiai tájfajtákat az introgresszív hibridizáció révén kihalás fenyegeti. Wiegand (1935) írta le először ezt a jelenséget növényeknél. Az introgresszió a gének taxonok közötti beszivárgását jelenti F1 hibridek hídján keresztül. E keresztezések termékeny utódai hibrid erőt (fokozott fitneszt) mutathatnak, és az egyik vagy mindkét szülői populációt kiszoríthatják (Ellstrand 2003). Azoknak a populációknak az újabb filogenetikai vizsgálatai, amelyek állítólag „Indica”-t és „Sativa”-t képviselnek, csekély vagy semmilyen genetikai különbséget nem mutatnak, mivel ezek a tanulmányok elsősorban hibrid „fajtavonalakat” elemeztek (Sawler et al. 2015; Dufresnes et al. 2017; Schwabe and McGlaughlin 2018). Ezek az eredmények ellentmondanak az 1970-es–1990-es években gyűjtött tájfajták vizsgálatainak, amelyek sokkal világosabb genetikai különbségeket mutattak (Hillig 2005a; Gilmore et al. 2007).
Az Indica–Sativa hibridek „strain” neveinek használata Watson (1985) munkájával kezdődött. Egy fajtavonalnév-adatbázis jelenleg 14 348 ilyen nevet sorol fel (Seedfinder 2019). Ez a közösségi alapon létrejött vállalkozás – nagyrészt titkos szülői hátterű hibridek keresztezése és újrakeresztezése – genetikai diverzitásvesztést eredményezett (Mudge et al. 2018). A vetőmagcégek által árult legtöbb fajtavonalat „Sativa-domináns” vagy „Indica-domináns” jelöléssel jellemzik. E megjelölések önkényességét mutatja az „AK-47”, egy hibrid fajtavonal, amely az 1999-es Cannabis Cup-on „Best Sativa” díjat nyert, négy évvel később pedig „Best Indica” díjat (McPartland 2017). Fogalmilag a „strain” egyenértékű a „cultivar”-ral; utóbbi a „International Code of Nomenclature for Cultivated Plants” (ICNCP, Brickell et al. 2016) által elismert taxonómiai rang. A drog-típusú Cannabis kevés kereskedelmi „strain”-je felelt meg azonban az ICNCP cultivar-elismerési követelményeinek (Small 2015).
Az ICNCP a cultivárokat „Groups” kategóriákba rendezi. Az ICNCP követelményeivel összhangban Small (2015) a közép-ázsiai tájfajtákat „Cannabis Group Narcotic, THC/CBD Balanced”, a dél-ázsiai tájfajtákat pedig „Cannabis Group Narcotic, THC Predominant” néven jelölte meg. Egyes botanikusok amellett érvelnek, hogy az emberi szelekcióval létrehozott bélyegekkel rendelkező növényeknek az ICNCP szerinti cultivar státuszt kellene adni, nem pedig taxonokat kellene hozzájuk rendelni a „International Code of Nomenclature for Algae, Fungi, and Plants” (ICN, Turland 2018) alapján. Gyakorlati okokból azonban a botanikusok az ICN keretrendszerét használják arra, hogy taxonokat rendeljenek mesterségesen szelektált növényekhez (pl. Hammer and Gladis 2014).
A fenti információk alapvetően a domesztikált anyagra vonatkoztak. Emellett a „vad” növények is figyelmet érdemelnek. A Cannabis „vad-típusú” bélyegeit először Zinger (1898) írta le: kis kaszatméret, tartós virágtakaró álcaszerű foltozottsággal, valamint megnyúlt alap – rövid, elkeskenyedő csonk formájában, jól fejlett leválási zónával. Ezzel szemben a domesztikált növények a vad-típusú növényekből hiányzó fenotípusos bélyegegyüttest (a „domesztikációs szindrómát”) fejeznek ki, például megnövekedett magméretet, a magpergés hiányát (a leválási zóna redukciója miatt) és a virágtakaró tapadásának csökkenését.
A domesztikált Cannabis könnyen kiszökik a termesztésből és „elvadul”. A domesztikált C. sativa Kanadában mindössze 50 generációval (évvel) a termesztés betiltása után visszatért vad-típusú fenotípushoz (Small 1975). Ez a gyors fenotípusos evolúció megnehezíti a valóban vad növények megkülönböztetését azoktól a korábban termesztett növényektől, amelyek visszatértek vad-típusú fenotípushoz. Így a termesztésen kívül növő Cannabis növények lehetnek (1) „önkéntesek” (a termesztésből nagyon nemrég kiszököttek, domesztikált jellemzőiket megtartva, és annak közelében növekedve, ahol termesztették őket); (2) „szökevények”, amelyek újra alkalmazkodtak a vad léthez (különféle élőhelyeken, tipikusan zavart vagy gyomos helyeken növekedve); vagy (3) „őshonosak/aboriginal” (domesztikáció által meg nem változtatottak, és őshonos területeiken növők).
A világ több legfontosabb kultúrnövényének őshonos populációi látszólag nem maradtak fenn, és a Cannabis is lehet ilyen természetű. Ettől függetlenül az Ázsiában vadon növő, a domesztikátumokhoz közeli (szimpatrikus vagy parapatrikus) növények különleges jelentőségűek. Lehetnek a domesztikátumok közvetlen ősei, bár ezt még tisztázni kell – sok ősi domesztikátumot a származási helyétől távoli helyeken domesztikáltak (Jarvis et al. 2016). Mindenesetre jelentős a valószínűsége annak, hogy a domesztikátumok közeli vad növényei géneket osztanak meg velük, mivel a Cannabis hatalmas mennyiségű virágport termel, amely nagy távolságokra terjed, és minden Cannabis-populáció képes keresztbeporzásra és teljesen interfertilis (Small 1972). Ennek megfelelően az ebben a közleményben elismert vad változatok igen jelentős potenciális génforrásokat képviselnek, amelyek a veszélyeztetett „Sativa” és „Indica” genomokat reprezentálják.
Ez a tanulmány nem tárgyalja az európai alfajt, a C. sativa subsp. sativa-t. Small and Cronquist (1976) ezt az alfajt két változatra osztották – domesztikált és vad-típusú növényekre. A domesztikált változat rosttípusú és olajmag-tájfajtákból, valamint cultivárokból áll. A vad-típusú változatnak nevezéktani problémái vannak a C. sativa var. spontanea Vavilov (1922) és a C. ruderalis (Janischevsky 1924) tekintetében. Vavilov és Janischevsky ezeket a külön taxonokat ugyanahhoz a vad-típusú növénypopulációhoz rendelték, amely az oroszországi Szaratov közelében nőtt. A „Ruderalis” a mai népi taxonómia alapelemévé vált (Anderson 1980). E nevezéktani kérdések megvitatását lásd a Kiegészítő anyag 1: SF.2 részében, a „vad-típusú nominalizmus” részletezését pedig az SF.3b-ben.
Az „Indica” és „Sativa” világszintű introgresszív hibridizációja veszélyezteti a C. sativa agrobiodiverzitását. Pesszimista nézőpontból az itt leírt változatok egy eltűnő világ alkotórészei, és osztályozásuk olyan, mint a dinoszauruszok újranevezésének gyakorlata. Optimista nézőpontból az őshonos közép- és dél-ázsiai változatok formális elismerése egyértelmű neveket ad nekik, és segíthet megakadályozni kihalásukat.
Módszerek
Az elemzéshez használt taxonómiai bélyegek a morfológia, fitokémia, genetika és gazda–parazita kapcsolatok szempontjait foglalták magukban. Egyes adatok újak (herbáriumi példányok morfológiai vizsgálatai), míg a fitokémiai és molekuláris adatokat korábban publikált tanulmányokból nyertük ki. E tanulmányok többsége közös kertkísérleteket (CGE-ket) alkalmazott. A CGE-k különböző helyekről származó növényeket egyetlen helyszínen, közös környezeti feltételek között, egységes feldolgozás mellett nevelnek (Grassi and McPartland 2017).
Morfológiai bélyegek
Körülbelül 1100 herbáriumi példányt vizsgáltunk meg 15 herbáriumban, amelyeket az Index Herbariorum herbáriumi akronimái jelölnek (Kiegészítő anyag 1: SF.4). Ezenfelül morfológiai adatokat nyertünk ki olyan CGE-kből, amelyek az előző században gyűjtött közép- és dél-ázsiai génanyagot hasonlították össze (pl. Vavilov and Bukinich 1929, Small et al. 1976, Anderson 1980, de Meijer 1994, Hillig 2005b). Archeobotanikai vizsgálatok morfológiai adataira is támaszkodtunk. A nyílt hozzáférés szellemében a kinyert morfológiai adatokat a Kiegészítő anyag 1: SF.8 részében közöljük, lehetővé téve, hogy az olvasók maguk szintetizálják a nyers adatokat. A CGE-vizsgálatok olyan adatokat szolgáltattak, amelyek gyakran hiányoznak a herbáriumi példányokból, például növénymagasságot, internódiumhosszt, szárvastagságot, valamint ágszöget vagy elágazási széttartást.
Az ágszög vagy elágazási széttartás azt a fokban mért szöget jelentette, amelyben egy ág a függőleges hajtásról levált; ez általában a függőlegestől számított 35° és 85° között mozgott. Az ágszög az internódiumhossz függvénye lehet, amelyet szintén értékeltünk. Az ághajlékonyság minőségi mérőszáma annak, hogy egy ág képes-e törés nélkül hajolni vagy lekonyulni. A hajlékonyság valószínűleg a háncsrost (rugalmas) és a farost (rugalmatlan) arányát tükrözi. A levélmorfológiát a virágzatok alapja közelében lévő „legyezőleveleken” (azaz nagyobb, tenyeresen összetett leveleken) értékeltük. A mintázott levelek megfeleltek a főhajtásról eredő elsőrendű elágazás fogalmának, ahogyan azt Spitzer-Rimon et al. (2019) bemutatták. A középső levélke hossz/szélesség aránya (L/W) hányadosként van kifejezve. A levélke alakja vagy lándzsás volt (a legszélesebb rész a levélke alapjától számított hossz felénél közelebb esik az alaphoz), vagy visszás-lándzsás (ahol a legszélesebb hely a hossz felénél távolabb van). Ezt a legszélesebb pontig mért távolság (WP) és a teljes hossz (WP/L) hányadosaként mértük. A WP/L > 0,5 értékű levélke visszás-lándzsás (Anderson 1980).
A perigoniális murvalevél (amelyet bracteolának, perigóniumnak, vagy helytelenül „csészének” is neveznek) az a virági murvalevél, amely körülzárja a nőivarú virágot, majd később a kaszatot (Small 2015). A virágzatsűrűséget minőségileg értékeltük a „perigoniális murvalevél–levél index” (azaz a „csésze–levél arány”, Clarke 1981) segítségével. Az alacsony indexű virágzatokban a levélanyag dominál – a fürtök között elhelyezkedő interszticiális „cukorlevelek” (viszonylag kis, kevés levélkéjű levelek a virágzatban), amelyek a 2. rendűtől a 7. rendűig terjedő oldalágacskákat támasztják alá (Spitzer-Rimon et al. 2019). Az alacsony index részben rövid internódiumhosszal és széles levélkeszélességgel társul.
A fejecskés-nyeles mirigyszőrök (CSGT-k) sűrűségét minőségileg értékeltük (azaz vizuálisan becsültük) a perigoniális murvaleveleken. A CSGT-sűrűséget Christison (1850) említette az egyik első olyan CGE-ben, amely a C. sativa-t (skót kendert) és a C. indica-t (indiai gunjuh-t) hasonlította össze. Megjegyezte, hogy a C. indica virágzatai érintésre gyantásak voltak: „A virágzati levelek, murvalevelek és virágtakaró mirigyes szőrözettel borítottak.” Azt is megjegyezte, hogy a C. indica levelei „ülő mirigyeket és mirigyszőröket [CSGT-ket] egyaránt” termeltek. A cukorleveleken lévő CSGT-sűrűséget szintén minőségileg értékeltük Potter (2009) módszere alapján.
Az itt használt értelemben a „termés” magában foglalja a kaszatot és annak többé-kevésbé tapadó virágtakaróját. A Cannabis nőivarú virágaiban a virágtakaró nem képez pártát, hanem az exokarpiumhoz (a kaszatfal legkülső rétegéhez) tapad. A termés méreteit és megjelenését értékeltük.
Minden herbáriumi példány esetében standardizált űrlapot használtunk a példánycímke adatainak (gyűjtő neve, dátum, hely, annotációk) és a morfológiai adatoknak a rögzítésére. A vizsgálat során morfológiai bélyegeket adtunk hozzá (pl. ágszög, virágzatsűrűség, CSGT-sűrűség), ami egyes herbáriumokba (BM, ECON, GH, IND, K) ismételt látogatásokat tett szükségessé. A morfológiai adatokat minőségileg (pl. ághajlékonyság, levélszín, virágzatsűrűség, CSGT-sűrűség, virágtakaró-tapadás), illetve mennyiségileg (pl. növénymagasság, internódiumhossz, levélke L/W és WP/L arányok, kaszatméret) szintetizáltuk. A mennyiségi adatok zárójelben megadott mérési tartományokat szolgáltattak minden leírt taxonhoz.
Fitokémiai bélyegek
Turner et al. (1980) széles körben idézett cikke 420, a C. sativa-ból izolált fitokemikáliát sorolt fel – a 420-as növényt. Kevés fitokemikália szolgáltat azonban hasznos taxonómiai információt. Vizsgálatunk a kannabinoidokra és terpenoidokra összpontosított. Élő növényekben és frissen betakarított szövetekben a kannabinoidok túlnyomórészt karbonsavak formájában vannak jelen. A THC tetrahidrokannabinolsavként (THCA) fordul elő; a kannabidiol (CBD) kannabidiolsavként (CBDA) fordul elő. A kannabinoidok dekarboxileződése semleges megfelelőikké öregedés során viszonylag lassan, hő hatására gyorsan megy végbe. Így a THCA THC-vé, a CBDA pedig CBD-vé alakul. Ezenkívül amikor a THC öregszik (ha nem megfelelően tárolják), jelentős mértékben kannabinollá (CBN), oxidációs termékké alakul. Ebben a cikkben, amikor THC-t és CBD-t említünk, a kontextustól függően úgy kell érteni, hogy a „THC” jelentheti a THCA + THC + CBN összességét, a „CBD” pedig a CBDA + CBD összességét.
A kannabinoidmennyiség (azaz THC% w/w) helyett a kannabinoidminőséget mérő paramétert közlünk: a THC/CBD arányt (THC% w/w osztva CBD% w/w-vel). A THC/CBD arány igen konzervatív (stabil) bélyeg, míg a THC% korrelál a morfológiával, például a trichomasűrűséggel (Potter 2009), valamint a virágzatsűrűséggel és a mirigyfejek méretével. Ezek a morfológiai különbségek nem változtatják meg a THC/CBD arányt. Az arányt egyetlen, kodomináns allélokkal rendelkező gén (de Meijer et al. 2003), vagy két szorosan kapcsolt, de különálló THCAS és CBDAS gén határozza meg (Van Bakel et al. 2011, Laverty et al. 2019). Weiblen et al. (2015) egyetlen kvantitatív tulajdonságlokuszt (QTL) azonosítottak, amely a THC/CBD aránnyal társult.
Ezzel szemben a THC% kifejeződése poligénes, és sok olyan gén módosítja, amely morfológiai különbségekhez járul hozzá. A környezeti tényezők (fényintenzitás, hőmérséklet, talajtápanyagok stb.) módosítják a THC%-ot, de sokkal kisebb hatással vannak a THC/CBD-re. Dimenzió nélküli arányként a THC/CBD érvényesebb összehasonlítást tesz lehetővé olyan sok tanulmány között, amelyek különböző körülmények között neveltek növényeket (Grassi and McPartland 2017).
A tetrahidrokannabivarin (THCV) és a kannabidivarin (CBDV) a THC és a CBD rövid oldalláncú C19 analógjai. A THCV-hez és CBDV-hez vezető bioszintetikus út korán, a kannabinoid-„csővezeték” rezorcinol oldalán ágazik el. Egyes kutatók a C19 analógokat hozzáadják a THC/CBD arányokhoz, THC+THCV/CBD+CBDV formában (pl. Turner et al. 1980). Itt a C19 analógok százalékos arányát (THCV%+CBDV%) külön bélyegként kezeljük.
A terpenoidok alkotják a Cannabis „illóolaját”. A terpenoidok közé tartoznak a szénhidrogén-terpének és azok oxigéntartalmú származékai, amelyek alkoholokat, étereket, aldehideket, ketonokat és észtereket képeznek. Illékonyak, és a növény jellegzetes szagát adják. Christison (1850) megjegyezte, hogy az indiai gunjuh balzsamos illatot bocsátott ki, amely hiányzott a skót kenderből. A dél-ázsiai tájfajták gyakran „herbális” vagy „édes” illatúak, míg a közép-ázsiai tájfajták csípős vagy „skunky” aromát adnak (Clarke 1981).
Genetikai bélyegek
A közép- és dél-ázsiai populációk – amelyeket az utóbbi időkben nem hibridizáltak jelentősen – molekuláris genetikai vizsgálatai számszerűen korlátozottak. Húsz évvel ezelőtt, amikor a hibridizálatlan tájfajták sokkal könnyebben hozzáférhetők voltak, a molekuláris módszerek tompa eszközök voltak. Ma teljes genomok DNS-szekvenciáját tudjuk dekódolni, de a hibridizálatlan biodiverzitás tartományának jó reprezentációja nem áll rendelkezésre elemzésre, bár Ázsiából valóban reprezentatív génanyag gyűjtése még mindig lehetséges lehet. A herbáriumok természetesen felbecsülhetetlen értékű régebbi példánytárak, de az ázsiai gyűjtemények viszonylag korlátozottak, és különböző okokból a kurátorok gyakran nem tudták engedélyezni a régebbi gyűjtemények mintavételét.
Egyes CGE-vizsgálatokhoz herbáriumi voucher példányokat helyeztek el (Small and Beckstead 1973; Turner et al. 1973, 1979; de Meijer et al. 1992; de Meijer 1994; Hillig 2004, 2005a; Hillig and Mahlberg 2004; Gilmore et al. 2007), amelyeket megvizsgáltunk a morfológiával való korrelációk megállapítására. Más fitokémiai és genetikai vizsgálatok esetében a hozzáférések földrajzi eredetéről szóló jelentésekre támaszkodtunk.
Eredmények
E cikk elektronikus változata Portable Document Formatban (PDF), ISSN- vagy ISBN-számmal rendelkező műként az ICN szerint publikált művet képvisel (Turland 2018). Ezért az e cikk elektronikus közlésében szereplő új nevek az ICN alapján már önmagában az elektronikus kiadásból hatályosan publikáltak. Az e munkában szereplő új neveket benyújtottuk az International Plant Names Indexhez (IPNI, http://www.ipni.org), ahonnan elérhetővé teszik őket a Global Names Index számára.
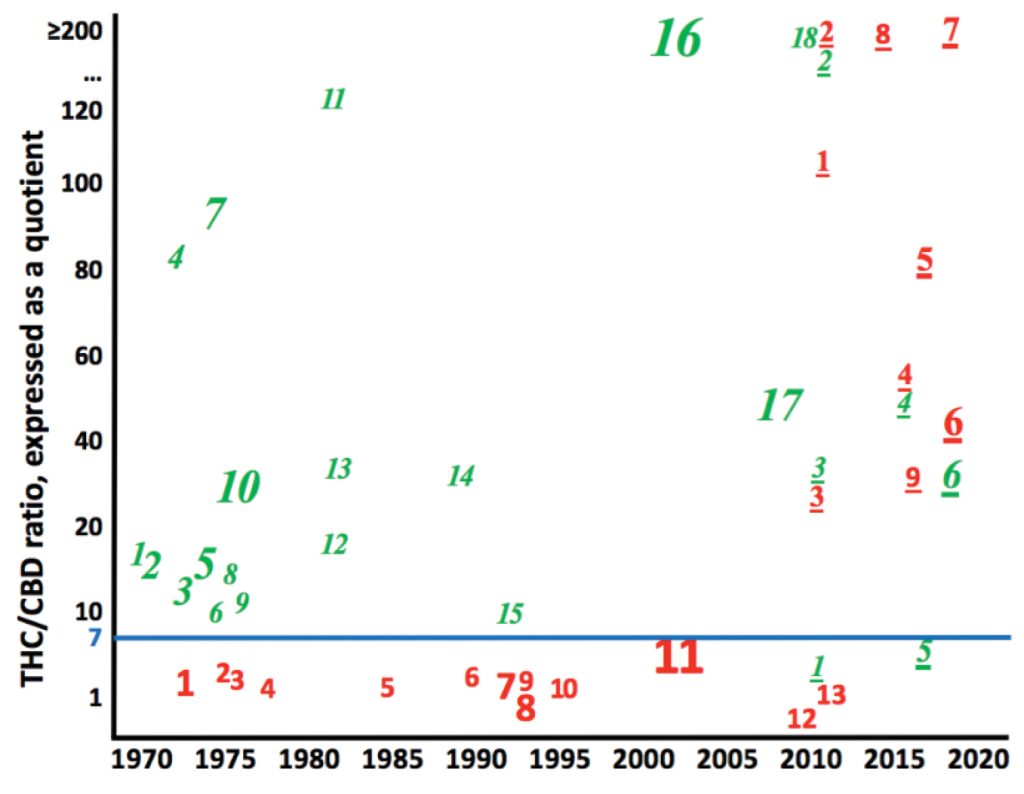
Az elmúlt 50 évben eltolódó taxonómiai bélyegre, ahogy a közép-ázsiai tájfajták hibridizálódtak az „Indica”-ba, a 2. ábra szolgáltat példát. A THC/CBD arányok elmúlt 50 évben bekövetkezett konvergenciáját szemlélteti. Az 1970-es–1990-es években gyűjtött hozzáférések vizsgálataiban a közép-ázsiai tájfajtákban (a dőlt nélküli piros betűtípusú tanulmányszámok) a THC/CBD arány, hányadosként kifejezve, mindig <7 volt (a tanulmányméret szerint súlyozott átlag = 3,56). Az 1970-es–1990-es években gyűjtött dél-ázsiai tájfajták vizsgálataiban (dőlt zöld betűtípusú tanulmányszámok) a THC/CBD arány ≥7 volt (a tanulmányméret szerint súlyozott átlag = 97,14). Azóta a THC/CBD arányok az állítólag Közép-Ázsiát képviselő hozzáférésekben (azaz „Indica”-ban) ugrásszerűen megnőttek. Ma már alig vagy egyáltalán nincs különbség az „Indica” és a „Sativa” között.
Taxonómiai elemzés
A C. sativa subsp. indica-t négy változatba soroltuk (formális nevezéktani értelemben, azaz varietas rangon). Két változat domesztikációs bélyegeket fejez ki (azonosak az „Indica” és „Sativa” kifejezések eredeti, szűk jelentésével), két változat pedig vad-típusú bélyegekkel rendelkezik. Követtük a Small and Cronquist (1976) által teremtett precedenst, akik a C. sativa subsp. indica-t két változatra – domesztikált és vad-típusú növényekre – osztották. Ezeket a változatokat azonban nem helyezték ős–utód kapcsolatba, mert a feltételezett ősi kapcsolatokat nem tudták igazolni.
Határozókulcs a C. sativa subsp. indica négy változatához¹
| 1. | A növények THC/CBD aránya rendszerint ≥7; a terpenoidprofil rendszerint nélkülözi a szeszkviterpén-alkoholokat, a friss aroma gyakran kellemes. A növények jó élőhelyeken ≥2 m magasak; az ágak hajlékonyak, a hajtásról viszonylag hegyes szögben (<45° a függőlegestől) térnek el. A friss levelek középzöld színűek; a középső levélkék keskenyek (hossz/szélesség általában >6), lándzsásak vagy lineáris-lándzsásak; a szélek finoman vagy durván fűrészesek, néha kétszeresen fűrészesek. Az érett nőivarú virágzat kissé kompakt (a virágzó szárak kis vagy közepes „bugákat”/„buds”-okat hoznak), viszonylag kevéssé feltűnő cukorlevelekkel (magas perigoniális murvalevél–levél index); a cukorleveleken a fejecskés-nyeles mirigyszőrök (CSGT-k) rendszerint a levelek proximális felére korlátozódnak; a perigoniális murvalevelek közepes vagy magas CSGT-sűrűséget mutatnak. Az érett kaszat exokarpiumának színe (a virágtakaró alatt) gyakran zöldesbarna. | |
| A | A THC/CBD arány mindig ≥7, gyakran sokkal magasabb. Az érett kaszatok rendszerint ≥3,6 mm hosszúak (3e, f ábra); a virágtakaró nagyrészt levált, de helyenként gyakran megmaradt (szabálytalan foltokként vagy csíkokként látható); a feltárt exokarpium erőteljes erezetet mutat; kiemelkedő, protuberáns alap hiányzik; a növényről nem válik le könnyen | var. indica („Sativa” történeti értelemben²). |
| B | A THC/CBD arány rendszerint ≥7, néha alacsonyabb. Az érett kaszatok rendszerint <3,6 mm hosszúak (3g, h ábra); a virágtakaró tartós (borítja az exokarpiumot és annak erezetét), erős, foltos vagy csíkos mintázatú pigmentációval; protuberáns alappal; a növényről könnyen leválik | var. himalayensis |
| 2. | A növények THC/CBD aránya <7; a terpenoidprofil szeszkviterpén-alkoholokat tartalmaz, a friss aroma gyakran csípős vagy „skunky”. A növények jó élőhelyeken <2 m magasak, és gyakran kb. 1 m-esek; az ágak nem hajlékonyak, az elágazás néha a szártengelytől közel 90°-os, menóra alakú habitust hozva létre. A friss levelek sötétzöldek, a nagyobb levelek levélkéi néha átfednek; a középső levélkék szélesek (hossz/szélesség általában <6), gyakran visszás-lándzsásak; a szélek durván fűrészesek, ritkán kétszeresen fűrészesek. Az érett nőivarú virágzat kompakt (a virágzó szárak közepes vagy nagy „bugákat”/„buds”-okat hoznak), feltűnő cukorlevelekkel (alacsony perigoniális murvalevél–levél index); a cukorleveleken a CSGT-k a levél hosszának több mint feléig húzódnak; a perigoniális murvaleveleket sűrű CSGT-borítás fedi. Az érett kaszat exokarpiumának színe (a virágtakaró alatt) gyakran világosabb olívazöldtől szürkéig terjed. | |
| A | A THC/CBD arány <7 (majdnem mindig >2). Az érett kaszatok rendszerint ≥3,6 mm hosszúak (3a, b ábra); a virágtakaró nagyrészt levált (szabálytalan foltokként vagy csíkokként látható); a feltárt exokarpium erőteljes erezetet mutat; kiemelkedő, protuberáns alap hiányzik; a növényről nem válik le, és gyakran a sűrű virágzatban reked | var. afghanica („Indica” történeti értelemben²). |
| B | A THC/CBD arány gyakran <2. Az érett kaszatok rendszerint <3,6 mm hosszúak (3c, d ábra); a virágtakaró tartós (borítja az exokarpiumot és annak erezetét), erős, foltos vagy csíkos mintázatú pigmentációval; protuberáns alappal; a növényről könnyen leválik | var. asperrima |
| ¹ Ahogyan a szöveg hangsúlyozza, az itt bemutatott különbségek hibridizálatlan növényeket képviselnek, a köztük végbement kiterjedt, újabb hibridizáció előtt. | ||
| ² Történetileg, amint a szöveg tárgyalja, a „Sativa” korábban a dél-ázsiai örökségű tájfajtákat, az „Indica” pedig a közép-ázsiai tájfajtákat képviselte. Ez a határozókulcs nem a ma kereskedelmi forgalomban kapható „Sativa” és „Indica” fajtavonalak azonosítására szolgál. | ||

Taxonómiai kezelés
Megjegyzendő, hogy a fényminőség herbáriumonként változott, ezért a herbáriumi példányokról és kaszatokról különböző herbáriumokban készült fényképek árnyalatban, színezetben és tónusban némileg eltértek. A négy változat protológusait (mindazt, ami egy bázionimmal a közlés idején társult) lásd a Kiegészítő anyag 1: SF.6 részében. A négy változat további reprezentatív herbáriumi példányaihoz lásd a Kiegészítő anyag 1-et.
1. változat: dél-ázsiai domesztikátum
Cannabis sativa subsp. indica var. indica (Lam.) Persoon, Synopsis Plantarum 2: 618, 1807.
4a ábra
Cannabis indica Lamarck, Encyclopédie Méthodique 1(2): 694–695, 1785. Bázionim. Persoonnak mint a comb. nov. szerzőjének idézésére vonatkozó indoklást lásd McPartland (1992) munkájában, és nem Wehmerét, ahogyan Small and Cronquist (1976) kezelték.
≡ C. sativa var. indica (Lam.) Fristedt, Upsala Läkareförenings Förhandlingar 5: 504, 1869–1870.
≡ C. sativa f. indica (Lam.) Voss in Siebert & Voss, Vilmorin’s Blumengärtnerei 1: 912, 1896.
≡ C. sativa var. indica (Lam.) Wehmer, Die Pflanzenstoffe p. 248, 1911.
= C. sativa var. indica Blume, Bijdragen tot de flora van Nederlandsch Indië, p. 515, 1825.
= C. macrosperma Stokes, Botanical Materia Medica 4: 539, 1812.
≡ C. sativa B macrosperma (Stokes) Ascherson & Graebner, Synopsis Mitteleuropäischen Flora 4: 599, 1911.
≡ C. sativa var. macrosperma (Stokes) Chevalier, Revue de Botanique Appliquée et d’Agriculture Coloniale 24: 64, 1944.
= C. sativa γ crispata Hasskarl, Neuer Schlüssel zu Rumph’s Herbarium amboinense p. 112, 1886.
= C. sativa β vulgaris de Candolle, Prodromus 16(1):31, 1869 (en part, indiai termesztett növények alapján).
= C. americana Houghton & Hamilton, Proc. Am. Pharm. Assoc. 55: 445, 1907, nomen nudum.
≡ C. americana Wehmer, Die Pflanzenstoffe, 2: 157, 1911, nomen nudum.
= C. madagascar Pearson, Proc. Penna. Pharm. Assoc. 1909: 179, 1909, nomen nudum.
= C. africana Glickman, Mulford’s Veterinary Bulletin 4(2): 88, 1912, nomen nudum.
≡ C. sativa var. africana Wehmer, Die Pflanzenstoffe 2: 39, 1935.
= C. mexicana Stanley, Am. J. Police Science 2(3): 252, 1931, nomen nudum.
Holotípus
India, valószínűleg Pondicherry, Lamarck, dátum nélkül, „Chanvre rapporte de l’Inde par M. Sonnerat” annotációval (herb. P). Pierre Sonnerat herb. P-ben lévő herbáriumi példányainak többségét Pondicherry környékén gyűjtötték 1775 és 1778 között.
Diagnózis
A növények virágzataiban a THC% ≥0,3%, és a THC/CBD arány mindig ≥7, gyakran jóval több; a középső levélke hossz:szélesség aránya ≥6 a virágzatok alapja közelében lévő legyezőlevelekben; az érett kaszatok rendszerint ≥3,6 mm hosszúak, a virágtakaró többnyire levált, hiányzik a kiemelkedő protuberáns alap, és hiányzik a jól fejlett leválási zóna, amely könnyű leválást tenne lehetővé.
Morfológia.
A növények rendszerint >2,0 m magasak (kedvezőtlen helyzetekben alacsonyabbak). A központi szár internódiumai viszonylag hosszúak (gyakran >12 cm, alacsonyabb növényekben rövidebbek), kissé üregesek (a szárátmérő legfeljebb 1/3-áig). Az ágak hajlékonyak, a szárról viszonylag hegyes szögben (kb. 45°) térnek el. A levél tenyeresen összetett, a legnagyobb levelek tipikusan legalább 7 levélkével rendelkeznek, a levélkeszélek nem fedik át egymást. A középső levélke hosszú és keskeny, lándzsás vagy lineáris-lándzsás alakú; a szélek közepesen durván fűrészesek, ritka másodlagos fűrészeltséggel. A nőivarú virágzat (és terméságazat) megnyúlt és kissé laza, viszonylag kevéssé feltűnő cukorlevelekkel (magas perigoniális murvalevél–levél index). A cukorleveleken a CSGT-k a proximális félre korlátozódnak. A perigoniális murvalevél közepes CSGT-sűrűséggel borított. A virágtakaró hártyás, hialin, pigmentált területekkel (barna, foltos vagy márványos megjelenésű); többnyire levált, de néha tartós. A kaszat rendszerint ≥3,6 mm hosszú, gömbölydedtől megnyúltig, exokarpiuma zöldesbarna; a leválási zóna gyengén fejlett.
Fitokémia
Szárított nőivarú virágzatok: THC ≥0,3%, a 20. század végi hozzáférésekben majdnem mindig >1,0%; szakirodalmi súlyozott x̄ = 3,97%, legfeljebb 12,5%. A THC/CBD arány ≥7, és gyakran >100. A THCV gyakran jelen van, különösen dél-ázsiai és afrikai tájfajtákban. Hillig and Mahlberg (2004) a THCV+CBDV% tartalomra x̄ = 0,25% értéket közölnek. A terpenoidprofil gyakran „herbális” vagy „édes” aromát ad, a terpinolén-, β-kariofillén-, transz-β-farnesén- és α-guaiene-tartalom pedig jelentősen magasabb, mint a közép-ázsiai növényekben.
Genetika
A dél-ázsiai örökségű tájfajták elkülönültek a közép-ázsiai tájfajtáktól egy alloenzim-elemzésben (Hillig 2005a) és egy cpDNS haplotípus-vizsgálatban (Gilmore et al. 2007). A „Sativa” és az „Indica” elkülönült STR-lókuszokkal (Knight et al. 2010), RAPD-markerekkel (Piluzza et al. 2013) és nDNS SNP-haplotípusokkal (Henry 2015; Lynch et al. 2016). Más vizsgálatok csekély vagy semmilyen genetikai különbséget nem mutattak ki a „Sativa” és „Indica” között (Sawler et al. 2015; Dufresnes et al. 2017), vagy fenotípusaik rosszul illeszkedtek állítólagos genotípusaikhoz (Schwabe and McGlaughlin 2018).
Egyéb bélyegek
Általában későn érő; a többi változathoz képest az egylaki növények viszonylag gyakoriak; fogékony a Schiffnerula cannabis által okozott fekete penészre.
Eredet és felhasználás. Eredetileg Indiában termesztették gañjā céljára, és korán elterjedt Délkelet-Ázsiába, Afrikába és Amerikába.

2. változat: dél-ázsiai vad-típus
Cannabis sativa subsp. indica var. himalayensis (Cazzuola) McPartl. & E.Small
4b ábra
Cannabis sativa var. hymalaiensis Cazzuola, Il Regno vegetale tessili e tintoriale, p. 49, 1875 (a helyesírás javítva az ICN 60.1 cikke szerint). Bázionim.
≡ C. sativa var. hymalaiensis Cazzuola, Nuovo Giornale Botanico Italiano 5: 262, 1873, nomen nudum.
≡ C. sativa var. himalayensis Cazzuola, Dizionario di botanica, p. 105, 1876 (későbbi homonima).
= C. sativa var. himalayensis Koch, Annales des Sciences Naturelles Botanique (Series 4) 1: 352, 1854, nomen nudum.
= C. sativa β vulgaris de Candolle, Prodromus 16(1):31, 1869 (en part, Észak-Indiában és Burmában spontán növő növények alapján).
= C. sativa α indica f. montana Fristedt, Upsala Läkareförenings Förhandlingar 5: 507, 1869–1870, nomen nudum.
= C. himalyana Zinger, Flora oder Allgemeine Botanische Zeitung 85: 207, 1898, nomen nudum.
= C. sativa subsp. indica sect. spontanea var. spontanea Clarke, Cannabis Evolution p. 224, 1987, nomen invalidum.
Neotípus
Itt kijelölve, INDIA: Himachal Pradesh, Shimla vagy Kinnaur („Himalaya Boreal. Occident., Regio Temp.”), T. Thompson, 1847 (GH). Cazzuola vagy Koch herbáriumaiban nem léteznek himalayensis példányok (személyes közlések, Lucia Amadei, herb. PI; Robert Vogt, herb. B). Thompson példányát neotípusként jelöltük ki, mert az általa a Himalájában készített több gyűjtés közül a legjobbat képviseli. Exsiccatumként terjesztették, több herbáriumban duplikátumokkal, izoneotípusokat szolgáltatva (BM! K! LE! US!).
Diagnózis
A növények virágzataiban a THC% ≥0,3%, és a THC/CBD arány gyakran ≥7, néha kevesebb; a középső levélke hossz:szélesség aránya ≥6 a virágzatok alapja közelében lévő legyezőlevelekben; az érett kaszatok rendszerint <3,6 mm hosszúak, tartós virágtakaróval és protuberáns alappal, és jól fejlett leválási zóna révén könnyen leválnak a növényről.
Morfológia
A növények 1,0–3,0 m magasak. A központi szár internódiumai viszonylag hosszúak (gyakran >10 cm, alacsonyabb növényekben rövidebbek), kissé üregesek (a szárátmérő legfeljebb 1/2-éig). Az ágak hajlékonyak, a szárról viszonylag hegyes szögben (kb. 45°) térnek el. A levél tenyeresen összetett, a nagyobb levelek rendszerint legalább 7 levélkével rendelkeznek, a levélkeszélek nem fedik át egymást. A középső levélke hosszú és keskeny, lándzsás alakú; a szélek közepesen durván fűrészesek, ritka másodlagos fűrészeltséggel. A nőivarú virágzat (és terméságazat) megnyúlt és kissé laza, viszonylag kevéssé feltűnő cukorlevelekkel (magas perigoniális murvalevél–levél index). A cukorleveleken a CSGT-k a proximális félre korlátozódnak. A perigoniális murvalevél közepes CSGT-sűrűséggel borított. A virágtakaró hártyás, hialin, pigmentált területekkel (barna, foltos vagy márványos megjelenésű); mindig tartós. A kaszat rendszerint <3,6 mm hosszú, exokarpiuma zöldesbarna; megnyúlt alappal és viszonylag keskeny leválási zónával.
Fitokémia
Szárított nőivarú virágzatok: THC ≥0,3% (bár két tanulmány THC <0,3%-ú növényekről számol be); súlyozott x̄ = 1,49%, tartomány 0,06% és 9,3% között. A THC/CBD arányok változnak; két tanulmány (azok, amelyek THC <0,3%-ot közöltek), közös hozzáféréseket vizsgálva, csak 1,28 és 1,56 arányt jelentett; ezek a hozzáférések olyan kelet-ázsiai rosttípusú domesztikátumokat képviselhetnek, amelyek visszanyerték a vad-típusú bélyegeket. Más tanulmányokban az arányok >10, sőt >100. A THC-tartalmat és THC/CBD arányokat torzítja a THCV%+CBDV%, amely magasabb, mint bármely más változatban: x̄ = 0,90% (Hillig and Mahlberg 2004). A terpenoidprofil hasonló a var. indica profiljához, kivéve a β-mircén, cisz-ocimén és β-kariofillén magasabb szintjét.
Genetika
Az alloenzim-elemzés (Hillig 2005a) részben elkülönítette a vad-típusú hozzáféréseket a dél-ázsiai domesztikátumoktól. Azt javasolta, hogy a Himalájából származó vad-típusú hozzáférések a dél-ázsiai domesztikátumok ősi forrását képviselik.
Egyéb bélyegek
Általában későn érő; a kaszatok éréskor lehullanak a növényről. A himalájai növények háncsrosttartalma (a szár száraz tömegének százalékában) magasabb, mint a kizárólag drogcélra Dél-Indiában termesztett növényeké (Bredemann 1952; de Meijer 1994).
Eredet és felhasználás
Vadon növő (lehetségesen őshonos) populációk fordulnak elő India hegyvidéki területein, Nepálban és Bhutánban, ahol háncsrostért (szárak), bhāngért (levelek), kézzel dörzsölt charasért (hashīsh) vagy kaszatokért (magokért) gyűjtik őket. Egyes himalájai herbáriumi példányok kaszatai viszonylag nagyok voltak, csökkent leválási mechanizmussal, ami domesztikált növényekből származó gének jelenlétére utal.
Bázionim-megjegyzések
Cazzuola 1873 és 1876 között többféleképpen írta a himalayensis epithetonját. Legkorábbi közlése nem adott világos diagnózist, nomen nudum volt, érvényesen nem publikálták (ICN Art. 38.2, Turland 2018). Koch is javasolt himalayensis taxont világos diagnózis nélkül, és azt a dél-ázsiai domesztikátummal azonosította – téves koncepcióként.
3. változat: közép-ázsiai domesztikátum
Cannabis sativa subsp. indica var. afghanica (Vavilov) McPartl. & E.Small, stat. nov.
urn:lsid:ipni.org:names:77208272-1
Cannabis sativa f. afghanica Vavilov, Trudy po Prikladnoi Botanike, Genetike i Selektsii 16(2): 227, 1926 (bázionim).
≡ C. indica var. afghanica Vavilov in Vavilov & Bukinich, Trudy Po Prikladnoi Botanike, Genetike i Selektsii 33 (Suppl.): 380, 1929, helyesírási változat.
≡ C. indica var. kafiristanica f. afghanica Vavilov in Vavilov & Bukinich, Trudy Po Prikladnoi Botanike, Genetike i Selektsii 33: 381, 1929.
= C. sativa subsp. culta prol. asiatica var. narcotica Serebriakova in Serebriakova & Sizov, Kul’turnaya Flora SSSR 5: 36, 1940 (latin diagnózis nélkül és tipifikálatlan).
= C. afghanica var. turkistanica Clarke, Cannabis Evolution p. 225, 1987, nomen invalidum.
= C. sativa var. afghanica McPartland, Hemp Diseases & Pests p. 4, 2000, nomen nudum.
= C. sativa var. afghan, Sands, U.S. patent 6,403,530, 2002, nomen nudum.
Neotípus
Itt kijelölve: Afganisztán: Ghazni tartomány (korábban Kandahar tartomány), Gui-Akhen (Гуй-Ахен) falu Qala-i Murvardar (Кала-и Мурвардар) közelében, a Ghazni–Kandahar úton, Vavilov, 1924, olyan magból, amelyet Serebriakova vetett 1926-ban a North Caucasus Experiment Station állomáson, Maikopban, Krasznodari határterületen (Cannabis sativa címkével, WIR 609, 3945). 5a ábra. A WIR-ben nem létezik afghanica címkéjű példány (McPartl., személyes megfigyelés, WIR 2010). A Vavilov and Bukinich (1929) által közölt kaszatábra nem szolgálhat lektotípusként, mert nem része a protológusnak, amely Vavilov (1926) munkájában jelenik meg.
Epitípus
Itt kijelölve, kifejezetten a neotípust támogatva: Afganisztán: Kandahar tartomány, Kandahar közelében, Schultes, XII.13–20.1971 (ECON 26505). 5b ábra. Az ICN az epitípust olyan példányként definiálja, amelyet értelmező típusként választanak ki, amikor a holo-/lekto-/neotípus kritikus azonosításra nem optimális (Turland 2018). Az ECON 26505 epitípusként szolgál, mert morfológiája egyértelműen egyezik az „Indica” széles körben elfogadott koncepciójával. Az ECON 26505 tipotípusként is szolgál – a példányról élő állapotban, a talajban készült fénykép megjelenik Schultes et al. (1974) munkájában, és a Kiegészítő anyag 1: SF.8 részében reprodukálták.
Diagnózis. A növények virágzataiban a THC% ≥0,3%, és a THC/CBD arány <7 (majdnem mindig >1); a középső levélke hossz:szélesség aránya <6 a virágzatok alapja közelében lévő legyezőlevelekben; az érett kaszatok rendszerint ≥3,6 mm hosszúak, a virágtakaró többnyire levált, hiányzik a kiemelkedő protuberáns alap, és hiányzik a jól fejlett leválási zóna, amely könnyű leválást tenne lehetővé.
Morfológia. A növények rendszerint <2 m magasak, gyakran <1 m. A központi szár internódiumai rövidek (gyakran 5–11 cm), többnyire tömörek, a központi üreg rendszerint a szárátmérő 20%-ánál kisebb. A jól fejlett növények ágai a talajszint közelében kezdődnek, néha a szártengelytől közel 90°-os szögben, menóra alakú habitust létrehozva. A levél tenyeresen összetett, a legnagyobb levelek tipikusan 7–11 levélkével, a levélkeszélek gyakran átfednek, a szín sötétzöld („fekete kender”, Vavilov 1992). A középső levélke hosszú és széles, gyakran visszás-lándzsás alakú; a szélek durván fűrészesek, másodlagos fűrészeltség ritkán látható. A nőivarú virágzat (és terméságazat) kompakt, gyakran trichomaváladékkal összeragadt, feltűnő cukorlevelekkel (alacsony perigoniális murvalevél–levél index); a rövid internódiumhossz miatt a hónalji fürtök összefolynak és tömött, kollektív colákká olvadnak össze. A cukorleveleken sűrű CSGT-k vannak a proximális félen, gyakran a levélke középpontján túl is jelen. A perigoniális murvalevelet sűrűn borítják CSGT-k. A virágtakaró hártyás, rendszerint levált, a termés alapja közelében csíkos vagy szabálytalanul foltos pigmentációs szegéllyel. A kaszat rendszerint ≥3,6 mm hosszú, exokarpiuma zöldtől szürkéig terjed; alapja tompa, jól fejlett leválási zóna nélkül.
Fitokémia
Szárított nőivarú virágzatok: THC ≥0,3, a 20. század végi hozzáférésekben majdnem mindig >1,0%; szakirodalmi súlyozott x̄ = 5,69%, legfeljebb 14,5%. Ez a változat fejezi ki az összes változat közül a legmagasabb teljes THC%+CBD%-ot (a növények relatív gyantatartalmának mérőszáma, mivel e két kannabinoid rendszerint dominálja a gyantát), ami korrelál a mirigyszőrök sűrű borításával. THCV%+CBDV% tartalma alacsonyabb, mint a dél-ázsiai populációké; Hillig and Mahlberg (2004) 0,14%-os átlagot közölnek. A terpenoidprofil csípős vagy „skunky” aromát ad, és egyedülállóan fejez ki szeszkviterpén-alkoholokat, például guaiolt, γ-eudezmolt, β-eudezmolt és a monoterpén-alkohol nerolidolt, valamint hidroxilált terpenoidokat, például γ-elemént, α-terpineolt és β-fencholt.
Genetika
Azokat az alloenzim- és DNS-vizsgálatokat, amelyek elkülönítették a közép-ázsiai és dél-ázsiai domesztikátumokat, az 1. változat genetikai szakasza részletezi. Onofri et al. (2015) egy SNP-t azonosítottak a THCA-szintázt kódoló génben, amely két afgán hozzáférésben és egy marokkói „hashīsh tájfajtában” volt egyedi (SNP-hozzáférési kódjuk: 1179, A→T transzverzió). Nem volt jelen 16 másik rost- és drog-típusú növényi hozzáférésben.
Egyéb bélyegek
Általában korai érésű, nagyobb késő szezonbeli fagytűréssel, mint a dél-ázsiai domesztikátumok. A késő szezonbeli hideg antociántermelést vált ki a levelekben és virágzatokban – ez a keresett „purple weed”. A kaszatok többnyire a növényeken maradnak, a sűrű terméságazat környező részei közé zárva. A növények fogékonyabbak a szürkerothadásra (Botrytis cinerea) és a lisztharmatra (Golovinomyces cichoracearum), mint a dél-ázsiai domesztikátumok.
Eredet és felhasználás
A 19. századból és a 20. század elejéről származó herbáriumi példányok Afganisztánból, Északnyugat-Pakisztánból, Turkesztánból (Üzbegisztán, Tádzsikisztán, Kirgizisztán, Xīnjiāng régió Kínában) és Iránból származnak. Ezeket a növényeket rostált hashīsh (nasha, charas) és néha magolaj céljára termesztették.
Megjegyzések
Vavilov (1926) az afghanica-t „a vad és termesztett kenderfajták közötti morfológiai kapocsnak” jellemezte. A Vavilov and Bukinich (1929) művében szereplő bizonyítékok azonban domesztikált fenotípust sugallnak (a Kiegészítő anyag 1: SF.6 részében érvelve). Small and Cronquist (1976) az afghanica-t domesztikátumként kezelték, a C. sativa subsp. indica var. indica alatt szinonimizálva. Small (2018) megjegyezte: „Az indica típusú marihuána jellemzői erősen összhangban vannak egy fejlett kultigén jellemzőivel. A modern olajmag-cultivárokhoz hasonlóan rövidek és kompaktak, olyan architektúrával, amely csökkenti az energia szártermelésbe való elterelését, és növeli a kívánt termék (virágzat) betakarítási indexét. Még a lombozat is (nagyon nagy, széles levélkékkel) összhangban van azzal a korábban leírt tendenciával, hogy a fejlett kultigének gyakran nagyobb leveleket mutatnak, mint vad és primitívebb termesztett rokonaik. Amikor az indica típusú fajtavonalakat magérlelésre hagyják (rendszerint virágzó anyagként takarítják be őket), a terméságazatok nagyon sűrűek, megakadályozva, hogy a magok többsége kihulljon és természetes módon terjedjen – ez a jelentős domesztikáció újabb jele.” A virágzatban lévő feltűnő cukorlevelek a domesztikáció másik jelét jelenthetik, mivel valószínűleg növelik a fejlődő virágokhoz és perigoniális murvaleveleikhez nagyon közel történő fotoszintetát-termelést.

4. változat: közép-ázsiai vad-típus
Cannabis sativa subsp. indica var. asperrima (Regel) McPartl. & E.Small
Cannabis sativa γ asperrima Regel, Acta Horti Petropolitani 6 (1): 476, 1879 (bázionim).
≡ C. sativa var. asperrima Regel in Herder, Acta Horti Petropolitani 12(1): 34, 1892.
= C. indica var. kafiristanica Vavilov in Vavilov & Bukinich, Trudy Po Prikladnoi Botanike, Genetike i Selektsii 33 (Suppl.): 381, 1929.
≡ C. sativa subsp. indica var. kafiristanica (Vavilov) Small & Cronquist, Taxon 24: 429, 1976.
≡ C. kafiristanica (Vavilov) Chrtek, Časopis Národního Muzea v Praze, Rada Přírodovědna 150(1–2): 22, 1981.
Lektotípus
Itt kijelölve: Kirgizisztán, Iszik-Kul régió, Karakol közelében, leg.: A. Regel; det.: E. Regel, 1.X.1877 (LE). 6a ábra.
Epitípus
Itt kijelölve, kifejezetten a neotípust támogatva: Afganisztán, Kunar tartomány, Chekhosarai (ma Asadābād), Vavilov, 1924, olyan magokból, amelyeket Serebriakova vetett 1927-ben a Pushkin Experiment Station állomáson, Detskoye Selo, Szentpétervár (WIR 599, 3952). 6b ábra.
Diagnózis
A növények virágzataiban a THC% ≥0,3%, és a THC/CBD arány <7 (majdnem mindig >1); a középső levélke hossz:szélesség aránya <6 a virágzatok alapja közelében lévő legyezőlevelekben; az érett kaszatok rendszerint <3,6 mm hosszúak, tartós virágtakaróval és protuberáns alappal, és jól fejlett leválási zóna révén könnyen leválnak a növényről.
Morfológia
A növények rendszerint <1,5 m magasak. A központi szár internódiumai rövidek (gyakran 5–11 cm, alacsonyabb növényekben rövidebbek), többnyire tömörek; a központi üreg, ha jelen van, rendszerint a szárátmérő 20%-ánál kisebb. A jól fejlett növények ágai a talajszint közelében kezdődnek, néha a szártengelytől közel 90°-os szögben, menóra alakú habitust hozva létre. A levél tenyeresen összetett, sötétzöld, a nagyobb levelek 5–7 levélkével rendelkeznek, néha átfednek. A középső levélke viszonylag rövid és széles, gyakran visszás-lándzsás alakú; a szélek durván fűrészesek, másodlagos fűrészeltség ritkán látható. A nőivarú virágzat kicsi, de kissé kompakt, mérsékelten feltűnő cukorlevelekkel (közepes perigoniális murvalevél–levél index). A cukorleveleken mérsékelten sűrű CSGT-k vannak a proximális félen. A perigoniális murvalevelet sűrűn borítják CSGT-k. A virágtakaró hártyás, sötétbarna pigmentációval foltos vagy néha lineáris mintázatban; tartós, de kézi manipulációval könnyen lepattogzik. A kaszat kicsi, oválistól megnyúltig, exokarpiuma sötét olívazöld színű, megnyúlt alappal.
Fitokémia
Szárított nőivarú virágzatok: THC ≥0,3, szakirodalmi súlyozott x̄ = 1,49%, tartomány 0,4% és 4,47% között. A THC/CBD arány szakirodalmi súlyozott x̄ = 2,23%, tartomány 0,77-től 4,75-ig (egy kiugró érték 9,43). A terpenoidprofil valószínűleg közelít a közép-ázsiai domesztikátuméhoz, de a szakirodalomban nem közölték.
Eredet és felhasználás
Az afghanica-ra hasonlító, de vad-típusú fenotípusú herbáriumi példányok eredete Északnyugat-Pakisztánból, Afganisztánból, Tádzsikisztánból, Üzbegisztánból, Kirgizisztánból, Kazahsztánból és a kínai Xīnjiāng régióból származik. E régió hegyei biodiverzitási „hotspotot” alkotnak, jelentős számú vad kultúrnövény-rokont és több mint 1000 endemikus növényfajt tartva fenn (Critical Ecosystem Partnership Fund 2017).
Megjegyzések
Herder (1892) a C. sativa γ asperrima-t külön változatként tartotta fenn, míg a C. erratica-t és a C. sativa β davurica-t a C. sativa alá szinonimizálta. E taxon közlési dátuma prioritást élvez Vavilov kafiristanica-jával szemben, de Vavilov példánya sokkal jobb állapotban őrződött meg, és epitípusként szolgál.

Megvitatás
A Cannabis populációi természetes és emberi szelekción egyaránt átestek. A fosszilis pollen-vizsgálatok azt mutatják, hogy a közép- és dél-ázsiai populációk legalább 32 600 éve elfoglalták külön ökológiai fülkéiket (McPartland et al. 2019). Fenotípusaik feltételezhetően divergáltak a környezeti adaptáció és természetes szelekció következtében. Közép-Ázsiára általában hűvösebb és szárazabb Köppen-klímák, valamint rövidebb tenyészidőszakok jellemzők. Dél-Ázsiára melegebb és nedvesebb Köppen-klímák, valamint hosszabb tenyészidőszakok jellemzők (Kottek et al. 2006).
A közép- és dél-ázsiai viszonyokhoz való ökológiai adaptációk valószínűleg élőhelyi izolációt, prezigotikus reprodukciós gátat hoztak létre. A dél-ázsiai viszonyok közé átültetett közép-ázsiai növények csökkent fitneszt (reproduktív sikert) szenvednek el. Amikor erősen virágzó ágaik monszunesőnek vannak kitéve, a terhelés alatt letörhetnek törékeny, menóra alakú elágazási habitusuk miatt. Ez nem fordul elő a dél-ázsiai növényekben, amelyek ágai hajlékonyabbak, és a szárról hegyesebb szögekben erednek. A közép-ázsiai növények sűrű, leveles virágzatai gyenge ellenállást mutatnak a nagy páratartalomban szaporodó gombákkal, például a Botrytis cinerea-val szemben. Ehhez képest a dél-ázsiai növények lazább, kevésbé leveles virágzatai jobban tűrik a nekrotróf gombákat (McPartland et al. 2000). A prezigotikus reprodukciós gátak további példáiért lásd a Kiegészítő anyag 1: SF.1 részét.
Az ArcGISPro 2.2 használatával feltérképeztük a vad-típusú var. asperrima-ként és var. himalayensis-ként azonosított herbáriumi példányok elterjedését (7. ábra). A himalayensis és asperrima herbáriumi példányok elterjedése összehasonlítható két korábbi közleménnyel, amelyek ezeket a földrajzi tartományokat térképezték: Indian Hemp Drugs Commission (1894) és Breckle and Koch (1982), reprodukálva a Kiegészítő anyag 1: SF.4 részében.
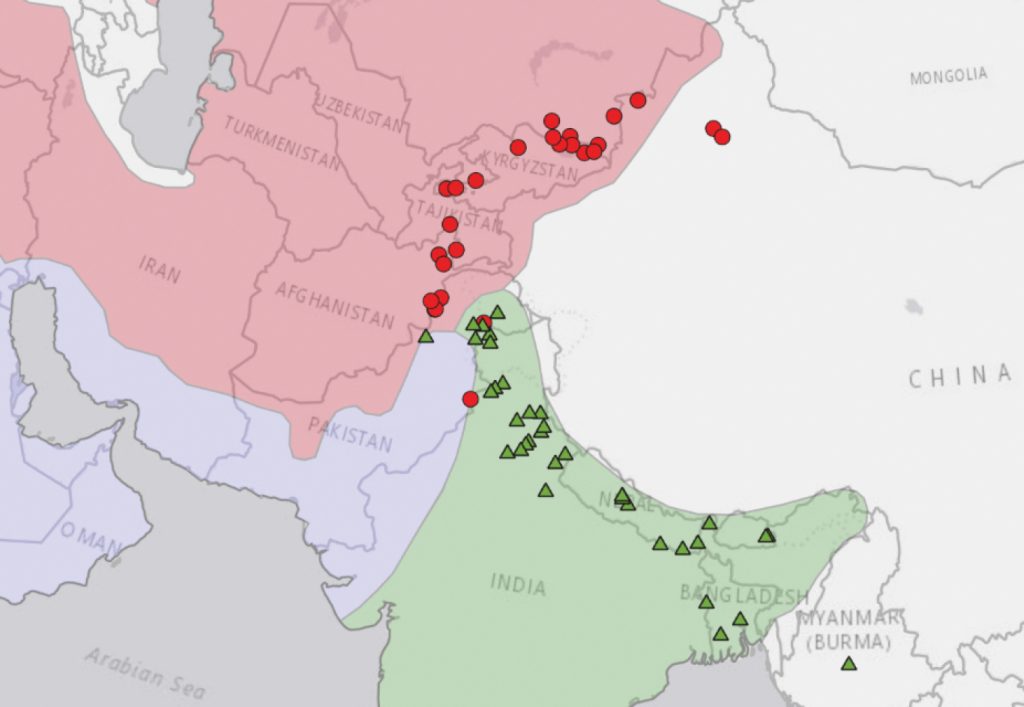
A himalayensis és asperrima elterjedése parapatrikus – areáik nem fednek át jelentősen, de egymás mellett helyezkednek el. Érintkezési felületük az Indus folyó vízgyűjtője (a var. himalayensis északnyugati határa) és a Kunar/Chitral folyó vízgyűjtője (a var. asperrima délkeleti határa) között fekszik. A parapatria támogatja az élőhelyi izolációról szóló hipotézisünket. A vad-típusú növények elterjedése hegyvonulatok ívén húzódik át Közép-Ázsiában (Hindu Kush, Karakoram, Pamír és Tien-san), valamint Dél-Ázsiában (Himalája és Purvanchal-hegység).
A közép- és dél-ázsiai eltérő klímák jellegzetes flórát eredményeznek, és a biogeográfusok Közép-Ázsiát és Dél-Ázsiát külön florisztikai régiókba sorolják. A florisztikai régiók a világ jól meghatározott területei, amelyeket viszonylag egységes növényfaj-összetételük, köztük endemikus flórájuk alapján ismernek fel. A 7. ábrán térképezett florisztikai régiók Djamali et al. (2012) alapján készültek. A var. asperrima herbáriumi példányai az iráni–turáni régióban lokalizálódnak, míg a var. himalayensis herbáriumi példányai az indiai régióban. Parapatrikus érintkezési felületük a szaharo–szindhi régióban fekszik. Más florisztikai régiókban lévő kiszórt előfordulások valószínűleg naturalizálódott szökevények herbáriumi példányait képviselik (korábban domesztikált növényeket, amelyek visszanyerték a vad-típusú bélyegeket).
Megjegyzendő, hogy Djamali et al. (2012) indiai florisztikai régiója White and Léonard (1991) munkájából frissített és egyszerűsített változat volt, akik a félszigeti Indiát elválasztották a Himalája vonulatától. Ők viszont Takhtajan (1986) munkáját egyszerűsítették, aki a Himalája vonulatát keleti és nyugati tartományokra osztotta, a nepáli Kali Gandakit tekintve választóvonalnak. Takhtajan az „Keleti Himalájai Tartományt” a Kínából érkező flóra beáramlása miatt különítette el. Feltételezzük, hogy ez volt az az útvonal, amelyen a Cannabis a Himalájába, majd onnan a félszigeti Indiába jutott. Viszonylag nemrég érkezett; Dél-Ázsia teljes területén a legidősebb fosszilis pollen mindössze 32 600 éves (McPartland et al. 2019). A var. himalayensis morfológiája közös bélyegeket mutat a kelet-ázsiai kenderrel, például magas termetet, viszonylag üreges hajtásokat magas háncsrost- és alacsony fatartalommal; levélkéket közepesen durva fűrészeltséggel; megnyúlt és kissé laza virágzatokat, magas perigoniális murvalevél–levél indexszel. A himalájai növények és a kelet-ázsiai kender hasonló THC/CBD arányokat (Kiegészítő anyag 1: Table S11) és terpenoidprofilokat (Kiegészítő anyag 1: Table S15) osztanak meg.
A korai földművelők a Cannabis evolúciójának következő körét indították el. A florisztikai régiók „diverzitási központokká” (COD-k) váltak, ahol a vad-típusú növényeket domesztikálták. Vavilov (1935) világszerte nyolc COD-t nevezett meg és térképezett fel. Előrelátóan két külön COD-t nevezett meg a Cannabis indica számára: a „közép-ázsiai COD”-t, amely az iráni–turáni florisztikai régiónak felel meg, és az „indiai COD”-t, amely az indiai florisztikai régiónak felel meg.
A közép- és dél-ázsiai populációk különböző emberi kezelési rendszerek alatt tovább divergáltak (amelyek szintén éghajlati szelekciónak voltak kitéve). A közép-ázsiaiak rostált hashīsh-t állítottak elő, ahol a tömeges feldolgozás valószínűleg korlátozta az egyedi magas THC-tartalmú növények szelekcióját (de Meijer 1999). Így a THC/CBD arányok közel maradtak a vad-típushoz. A dél-ázsiaiak gañjā-t állítottak elő, ahol a növényeket egyedileg lehetett betakarítani, és a dél-ázsiaiak a kiválasztott, magas THC-tartalmú növények magjait szelektálták, ezáltal egy évezred folyamán növelve a THC/CBD arányokat (Clarke and Merlin 2013).
A dél-ázsiai génanyagot a 13. századra Délkelet-Ázsiába és Kelet-Afrikába, majd az afrikai rabszolga-kereskedelem során Brazíliába vitték (Clarke and Merlin 2013). A közép-ázsiai domesztikátum az 1970-es évek előtt korlátozott elterjedésű volt, Afganisztánra, Pakisztánra és Turkesztánra korlátozódva. A turkesztáni növényeket néha dél-ázsiai domesztikátumokként osztályozzák (Clarke and Merlin 2013; Small 2015), bár Clarke (1987) a turkesztáni domesztikátumokra C. afghanica var. turkistanica [sic] taxont állított fel. A 19. századi herbáriumi gyűjtemények azt jelzik, hogy a termesztett turkesztáni növények közép-ázsiai domesztikátumok voltak, nem dél-ázsiaiak.
E vizsgálat célja az volt, hogy „gyakorlati és természetes” taxonokat azonosítson a C. sativa subsp. indica-n belül. Döntésünk, hogy az alfajt négy változatra hasítjuk, vitákat vet fel a nevezéktani prioritásokról, beágyazott hierarchiákról és gyakorlati alkalmazásokról. Ezeket a kérdéseket a Kiegészítő anyag 1: SF.13 részében tárgyaljuk. Hangsúlyunk a domesztikátumokon volt, amelyek a dél-ázsiai örökségű tájfajtákat (C. sativa subsp. indica var. indica) és a közép-ázsiai tájfajtákat (C. sativa subsp. indica var. afghanica) képviselik. Több bélyeg hajlamos elkülöníteni ezeket a taxonokat (1. táblázat). Legjobban THC/CBD arányaik és terpenoidprofiljaik alapján különíthetők el.
- táblázat. A háziasított, magas THC-tartalmú változatok, a C. sativa subsp. indica var. indica és a C. sativa subsp. indica var. afghanica megkülönböztetésére szolgáló trendek.¹
|
Jellemző |
C. s. var. indica |
C. s. var. afghanica |
|
THC/CBD arány |
≥7 |
<7 |
|
THCV+CBDV tartalom |
Gyakran jelen |
Gyakran hiányzik |
|
Terpenoidprofil |
„Herbális” vagy „édes” aroma, szeszkviterpén-alkoholok nélkül |
Csípős vagy „skunky” aroma, guaiol, γ-eudezmol és β-eudezmol jelenlétével |
|
Magasság, elágazás |
A jól fejlett növények rendszerint ≥2 m magasak; az elágazás hajlékony (felfelé irányuló habitussal) |
A jól fejlett növények rendszerint <2 m magasak; az elágazás rugalmatlan (menóra alakú habitussal) |
|
Levelek a virágzatok alapján |
Világosabb zöld, rendszerint 7 levélkével, a levélkeszélek között hézagokkal |
Sötétebb zöld, rendszerint 9 levélkével, átfedő szélekkel |
|
Több levélkéjű levelek középső levélkéi |
Hosszúak és keskenyek, lándzsás vagy lineáris-lándzsás alakúak; a szélek finoman fűrészesek, néha kétszeresen fűrészes szélek láthatók |
Hosszúak és szélesek, gyakran visszás-lándzsás alakúak; a szélek durván fűrészesek, kétszeresen fűrészes szélek ritkán láthatók |
|
Pisztillátus virágzatok |
Viszonylag lazák és nyíltak, a cukorlevelek viszonylag kevéssé feltűnők (magas perigoniális murvalevél–levél index) |
Kompaktak és feltűnő cukorlevelekkel rendelkeznek (alacsony perigoniális murvalevél–levél index) |
|
Nyeles mirigyszőr-sűrűség |
Kevés a virágzati levelek proximális végén; közepesen sűrű a perigoniális murvaleveleken |
Sok a virágzati levelek proximális végén, legalább a virágzati levelek feléig terjed; nagyon sűrű a perigoniális murvaleveleken |
|
Virágtakaró |
Foltos pigmentációjú virágtakaró, néha az egész kaszaton tartós |
Foltos pigmentációjú virágtakaró, ritkán tartós, a kaszat alapjára korlátozódik |
|
Kaszat |
Az exokarpium színe zöldesbarna (sötétebb, mint az afghanica-é), mérettartományának alsó része kisebb, mint az afghanica-é; lazán ágyazódik a perigoniális murvalevélbe és cukorlevelekbe |
Az exokarpium színe olívazöldtől szürkéig (világosabb, mint az indica-é), mérettartományának felső része nagyobb, mint az indica-é; szorosan ágyazódik a perigoniális murvalevélbe és cukorlevelekbe |
|
Érési idő |
Később érő |
Korábban érő |
|
Egyéb bélyegek |
Fogékony a fekete penészre (Schiffnerula cannabis), egylaki növények alkalmanként láthatók |
Fogékony a szürkerothadásra (Botrytis cinerea) és a lisztharmatra (Golovinomyces cichoracearum), egylaki növények ritkán láthatók |
¹ Ahogyan a szöveg hangsúlyozza, az itt bemutatott különbségek az „Indica” és „Sativa” tájfajták történeti, hibridizálatlan formáit képviselik, a köztük végbement kiterjedt, újabb hibridizáció előtt.
Az 1. táblázatban szereplő, a tájfajtákat megkülönböztető trendek közül kevés maradt igaz a mai kereskedelemben kapható „Indica” és „Sativa” fajtavonalakra. Különösen a THC/CBD arányok konvergáltak az állítólag „Indica”-t és „Sativa”-t képviselő anyagban (2. ábra). Az „Indica” és „Sativa” néhány újabb vizsgálata megfordulásokat mutat tájfajta őseikhez képest. Míg a közép-ázsiai tájfajták alacsonyabb THC/CBD arányokat fejeztek ki, mint a dél-ázsiai tájfajták, hat újabb tanulmány ennek fordítottját jelentette az „Indica” és „Sativa” esetében (Fischedick et al. 2010; Hazekamp and Fischedick 2012; Elzinga et al. 2015; Hazekamp et al. 2016; Lynch et al. 2016; Jikomes and Zoorob 2018). Ez arra késztette Hazekamp and Fischedick (2012) szerzőket, hogy elhagyják az „Indica”/„Sativa” nevezéktant a „chemovarok” javára.
Meglepő módon a terpenoidprofilok nagyrészt különállók maradtak. Az „Indica” hibridek, közép-ázsiai őseikhez hasonlóan, egyedülállóan fejeznek ki szeszkviterpén-alkoholokat. Ezek hiányoznak a dél-ázsiai tájfajtákból és „Sativa” leszármazottaikból (Kiegészítő anyag 1: SF.9). A THC-tartalomra irányuló évszázados mesterséges szelekció láthatóan nem változtatta meg a szeszkviterpén-alkohol-tartalmat. Ugyanez igaz lehet a THCV-re. Korlátozott bizonyítékok arra utalnak, hogy a THCV, amely a dél-ázsiai tájfajták és dél-ázsiai vad-típusok markere (Hillig and Mahlberg 2004), megmarad a „Sativa”-ban (Hazekamp and Fischedick 2012; Aizpurua-Olazizolo et al. 2016).
A változatok között gyakran figyelhetők meg köztes formák, amelyek a biológiai fajfogalom szerint képesek kereszteződni és géneket cserélni. Ahol a változatok földrajzilag átfednek, gyakran hoznak létre köztes formákat. A köztes formák gyakran láthatók pakisztáni herbáriumi példányokban, amely a subsp. indica diverzitási központja – mind a négy változat előfordul ott. A Közel-Keletről (Törökország, Szíria, Libanon, Palesztina, Izrael, Jordánia, Irak, Nyugat-Irán) és Észak-Afrikából (Egyiptomtól Marokkóig) származó számos herbáriumi példány szintén köztes fenotípusokat mutat. Clarke and Merlin (2013) a közel-keleti és észak-afrikai populációkat a dél-ázsiai tájfajták őseiként osztályozták. Közép-ázsiai génanyag azonban a 1200-as években, majd ismét az 1600-as években eljuthatott a Közel-Keletre (Kiegészítő anyag 1: SF.11).
Számos mennyiségi fenotípusos bélyeg mérése még várat magára a Cannabis-ban, például a mirigyszőrök sűrűsége mm² felületre vetítve, a mirigyszőrök mérete és a mirigyfejek leválása. A C. indica és C. afghanica megkülönböztetésére alkalmas egyértelmű genetikai „vonalkód” felfedezésre vár. Lásd a „Future directions” részt a Kiegészítő anyag 1: SF.13 részében. Végül, ez a tanulmány nem tárgyalta a kelet-ázsiai kendert. Kannabinoid- és genetikai adatok a kelet-ázsiai Cannabis-t a C. indica subsp. indica génkészletének részhalmazaként különítik el (Hillig 2005b). A kelet-ázsiai Cannabis-ról, különösen a yúnnáni biodiverzitásról bővebben lásd a Kiegészítő anyag 1: SF.12 részét.
Következtetések
Az itt körülhatárolt és megnevezett négy Cannabis-változat formális elismerést érdemel. Az infraspecifikus taxonok elismerése segít azonosítani a kihalásnak kitett populációkat (pl. Ellstrand 2003; Haig et al. 2006). Az Egyesült Nemzetek Biodiverzitási Egyezménye nyomán az infraspecifikus variáció a megőrzési erőfeszítések fókuszába került (Coates et al. 2018). A négy Cannabis-változat és egyedi morfológiai, valamint kémiai bélyegeik elismerése „prior art”-ot is szolgáltat, megakadályozva az eredetiségre vonatkozó igényeket a Cannabis hasznossági szabadalmaiban.
Sürgősen szükség van a közép- és dél-ázsiai tájfajták őshonos populációiból származó génanyag gyűjtésére és megőrzésére saját diverzitási központjaikban. A diverzitási központjaikon kívüli génanyagbázis a széles körű keresztezés miatt genetikailag szennyeződött. Az éghajlatváltozás és az előre nem jelezhető jövőbeli szükségletek kontextusában az agrobiodiverzitás in situ megőrzése sokkal előnyösebb a kultúrnövények és vad rokonaik számára, de tekintettel a Cannabis Ázsiában fennmaradt, megváltozatlan őshonos vad populációinak bizonytalan további létezésére, a magbankokban történő megőrzés azonnali prioritás. Remélhetőleg az itt adott egyértelmű nevek segíthetnek megakadályozni e taxonok kihalását.
Köszönetnyilvánítás
Karl Hillignek és Patty Pruittnak köszönjük az évtizedeken át nyújtott tanulságos meglátásokat. William Hegman ArcGIS térképezési szakértelmet biztosított. Köszönetet mondunk minden herbárium kurátorainak a példányokhoz való hozzáférésért, különösen Tamara Smekalovának és Zhuk Mikhailnak a WIR-ben, Walter Kittredge-nek az ECON-ban és Eric Knoxnak az IND-ben. E kutatást részben a GW Pharmaceuticals támogatta.
Tanulmány-információk
Eredeti közlemény
McPartland JM, Small E (2020) A classification of endangered high-THC cannabis (Cannabis sativa subsp. indica) domesticates and their wild relatives. PhytoKeys 144: 81–112. DOI: 10.3897/phytokeys.144.46700
Szerzői információk
Szerzők: John M. McPartland‡§, Ernest Small|
Intézményi hovatartozások: ‡ GW Pharmaceuticals, Cambridge, Egyesült Királyság; § University of Vermont, Burlington, Amerikai Egyesült Államok; | Agriculture and Agri-Food Canada, Ottawa, Kanada
Levelező szerző: John M. McPartland (mcpruitt@myfairpoint.net)
Tudományos szerkesztő: Hugo de Boer
Előzménydátumok
Beérkezett: 2019. szeptember 19.; elfogadva: 2020. február 2.; megjelent: 2020. április 3.
Szerzői jog és licenc
© 2020 John M. McPartland, Ernest Small. Ez nyílt hozzáférésű cikk, amely a Creative Commons Attribution License (CC BY 4.0) feltételei szerint terjeszthető; ez korlátlan felhasználást, terjesztést és bármely médiumban történő reprodukciót engedélyez, feltéve, hogy az eredeti szerzőt és forrást feltüntetik.
Adat-, kód- és kiegészítő anyagok
Kiegészítő anyag
Cím: A veszélyeztetett, magas THC-tartalmú kannabisz (Cannabis sativa subsp. indica) domesztikátumainak és vad rokonainak osztályozása
Szerzők: John M. McPartland, Ernest Small
Adattípus: fajadatok
Magyarázó megjegyzés: fajfogalmak, szintnominalizmus, vad-típusú nominalizmus, protológusok, nevezéktani prioritás, köztes formák és a módszertan részletezése. A morfológiai és a teljes bizonyítékokon alapuló elemzésekben használt taxonómiai bélyegek listája és azok kódolása.
Adatlicenc: Ez az adatkészlet az Open Database License (ODbL) alatt érhető el.
Ez az adatkészlet az Open Database License (http://opendatacommons.org/licenses/odbl/1.0/) alatt érhető el. Az Open Database License (ODbL) olyan licencmegállapodás, amelynek célja, hogy a felhasználók szabadon megoszthassák, módosíthassák és használhassák ezt az adatkészletet, miközben ugyanezt a szabadságot mások számára is fenntartják, feltéve, hogy az eredeti forrást és szerző(ke)t feltüntetik.
Fájl letöltése (7,83 MB)
Hivatkozások
Aizpurua-Olazizolo O, Sodaner U, Öztürk E, Schibano D, Simsir Y et al. (2016) Evolution of the cannabinoid and terpene content during the growth of Cannabis sativa plants from different chemotypes. Journal of Natural Products 79 (2): 324–331. https://doi.org/10.1021/acs.jnatprod.5b00949
Anderson LC (1980) Leaf variation among Cannabis species from a controlled garden. Harvard University Botanical Museum Leaflets 28 (1): 61–69.
Barbaro E (1516) In hoc volumin haec continentur, Ioannis Baptistae Egnatii in Dioscoridem ab Hermolao Barbaro tralatum annotamenta. Aloisius & Franciscus Barbari, Venice.
Beisler J (2006) The bandit of Kabul. Regent Press, Oakland, CA.
Breckle SW, Koch M (1982) Afghanische Drogen und ihre Stammpflanzen III. Haschisch und hanf. Afghanistan Journal 9 : 115–123.
Bredemann G (1952) Weitere Beobachtungen bei Züchtung des Hanfes auf Fasergehalt. Der Züchter 22 : 257–269. https://doi.org/10.1007/BF00710361
Brickell CD, Alexander C, Cubey JJ, David JC, Hoffman MHA et al. (Eds) (2016) International code of nomenclature for cultivated plants. Ninth edition. International Society for Horticultural Science, Leuven, Belgium.
Casano S (2005) Ottimizzazione dell’ agrotecnica della canapa ( Cannabis sativa L.) per applicazioni di tipo farmaceutico. Doctoral thesis, Università degli studi di Palermo.
Cherniak L (1982) The great books of Cannabis , Vol. I: Book II. Cherniak/Damele Publishing Co., Oakland CA.
Christison A (1850) On Cannabis indica , indian hemp. Transactions and Proceedings of the Botanical Society of Edinburgh 4 : 59–69. https://doi.org/10.1080/03746605309467592
Clarke RC (1981) Marijuana botany. And/Or Press, Berkeley, CA.
Clarke RC (1987) Cannabis evolution. MS thesis, Indiana University, Bloomington, IN.
Clarke RC (1998) Hashish! Red Eye Press, Los Angeles, CA.
Clarke RC, Merlin MD (2013) Cannabis evolution and ethnobotany. University of California Press, Berkeley.
Clarke RC, Merlin MD (2016) Cannabis domestication, breeding history, present-day genetic diversity, and future prospects. Critical Reviews in Plant Sciences 35 : 293–327. https://doi.org/10.1080/07352689.2016.1267498
Coates DJ, Byrne M, Moritz C (2018) Genetic diversity and conservation units: Dealing with the species–population continuum in the age of genomics. Frontiers in Ecology and Evolution 6: 165. https://doi.org/10.3389/fevo.2018.00165
Critical Ecosystem Partnership Fund (2017) Mountains of Central Asia biodiversity hotspot. https://www.cepf.net/our-work/biodiversity-hotspots/mountains-central-asia
de Meijer EPM (1994) Diversity in Cannabis . Doctoral thesis, Wageningen Agricultural University, Wageningen, The Netherlands.
de Meijer EPM (1999) Cannabis germplasm resources. In: Ranalli P (Ed.) Advances in Hemp Research. Haworth Press, New York , 133–151.
de Meijer EPM, van Soest LJM (1992) The CPRS Cannabis germplasm collection. Euphytica 62 (3): 201–211. https://doi.org/10.1007/BF00041754
de Meijer EPM, van der Kamp HJ, van Eeuwijk FA (1992) Characterization of Cannabis accessions with regard to cannabinoid content in relation to other plant characters. Euphytica 62 (3): 187–200. https://doi.org/10.1007/BF00041753
de Meijer EPM, Bagatta M, Carboni A, Crucitti P, Cristiana Moliterni VM et al. (two additional authors) (2003) The inheritance of chemical phenotype in Cannabis sativa L. Genetics 163 : 335–346.
Djamali M, Brewer S, Breckle SW, Jackson ST (2012) Climatic determinism in phytogeographic regionalization – A test from the Irano-Turanian region SW and Central Asia. Flora 207 (4): 237–249. https://doi.org/10.1016/j.flora.2012.01.009
Dufresnes C, Jan C, Bienert F, Goudet J, Fumagalli L (2017) Broad-scale genetic diversity of Cannabis for forensic applications. PLoS One 12(1): e0170522. https://doi.org/10.1371/journal.pone.0170522
Ellstrand NC (2003) Dangerous liaisons? When cultivated plants mate with their wild relatives. Johns Hopkins University Press, Baltimore.
Elzinga S, Fishchedick J, Podkolinski R, Raber JC (2015) Cannabinoids and terpenes as chemotaxonomic markers in Cannabis . Natural Products Chemistry & Research 3: 4. https://doi.org/10.4172/2329-6836.1000181
Fischedick JT, Hazekamp A, Erkelens T, Choi YH, Verpoorte R (2010) Metabolic fingerprinting of Cannabis sativa L., cannabinoids and terpenoids for chemotaxonomic and drug standardization purposes. Phytochemistry 71 (17–18): 2058–2073. https://doi.org/10.1016/j.phytochem.2010.10.001
Gilmore S, Peakall R, Robertson J (2007) Organelle DNA haplotypes reflect crop-use characteristics and geographic origins of Cannabis sativa . Forensic Science International 172 (2–3): 179–190. https://doi.org/10.1016/j.forsciint.2006.10.025
Gould J (2015) The Cannabis crop. Nature 525(7570): S2–S3. https://doi.org/10.1038/525S2a
Grassa CJ, Wegner JP, Dabney C, Poplawski SH, Motley ST, Michael TP, Schwartz CJ, Weiblen GD (2018) BioRxiv Preprint, https://doi.org/10.1101/458083
Grassi G, McPartland JM (2017) Chemical and morphological phenotypes in breeding of Cannabis sativa L. In: Chandra S, Lata H, ElSohly MA (Eds) Cannabis sativa : Botany and Biotechnology. Springer International Publishing, Cham, Switzerland , 137–160. https://doi.org/10.1007/978-3-319-54564-6_6
Haig SM, Beever EA, Chambers SM, Draheim HM, Buggar BD et al. (2006) Taxonomic considerations in listing subspecies under the U.S. endangered species act. Conservation Biology 6 (6): 1584–1594. https://doi.org/10.1111/j.1523-1739.2006.00530.x
Hammer K, Gladis T (2014) Notes on infraspecific nomenclature and classifications of cultivated plants in Compositae , Cruciferae , Cucurbitaceae , Gramineae (with a remark on Triticum dicoccon Schrank) and Leguminosae . Genetic Resources and Crop Evolution 61 (8): 1455–1467. https://doi.org/10.1007/s10722-014-0148-8
Hazekamp A, Fischedick JT (2012) Cannabis – From cultivar to chemovar. Drug Testing and Analysis 4 (7–8): 660–667. https://doi.org/10.1002/dta.407
Hazekamp A, Tejkalová K, Papadimitriou S (2016) Cannabis : From cultivar to chemovar II – a metabolomics approach to Cannabis classification. Cannabis and Cannabinoid Research 1 (1): 202–215. https://doi.org/10.1089/can.2016.0017
Hebert PD, Stoeckle MY, Zemlak TS, Francis CM (2004) Identification of birds through DNA barcodes. PLoS Biology 2(10): e312. https://doi.org/10.1371/journal.pbio.0020312
Henry P (2015) Genome-wide analyses reveal clustering in Cannabis cultivars: the ancient domestication trilogy of a panacea. PeerJ PrePrints. https://doi.org/10.7287/peerj.preprints.1553v2
Herder FG (1892) Plantae Raddeanae apetalae V. Acta Horti Petropolitani 12 (1): 31–132.
Herodotus (2007) The landmark Herodotus: the Histories. Pantheon Books, NY.
Hey J, Pinho C (2012) Population genetics and objectivity in species diagnosis. Evolution 66 (5): 1413–1429. https://doi.org/10.1111/j.1558-5646.2011.01542.x
Hillig KW (2004) A chemotaxonomic analysis of terpenoid variation in Cannabis . Biochemical Systematics and Ecology 32 (10): 875–891. https://doi.org/10.1016/j.bse.2004.04.004
Hillig KW (2005a) Genetic evidence for speciation in Cannabis ( Cannabaceae ). Genetic Resources and Crop Evolution 52 (2): 161–180. https://doi.org/10.1007/s10722-003-4452-y
Hillig KH (2005b) A systematic investigation of Cannabis . Doctoral Dissertation, Indiana University, Bloomington, IN.
Hillig KW, Mahlberg PG (2004) A chemotaxonomic analysis of cannabinoid variation in Cannabis ( Cannabaceae ). American Journal of Botany 91 (6): 966–975. https://doi.org/10.3732/ajb.91.6.966
Indian Hemp Drugs Commission (1894) Report of the Indian Hemp Drugs Commission, 1893–94. Government Central Printing Office, Simla.
Janischevsky DE (1924) Форма конопли на сорных местах в Юго-Восточной России. Ученые записки Саратовского государственного университета имени Н. Г. Чернышевского 2 (2): 3–17.
Jarvis DI, Hodgkin T, Brown AHD, Tuxill J, Lopez v I et al. (2016) The origins of agriculture, crop domestication, and centers of diversity. In: Jarvis DI et al. (Eds) Crop genetic diversity in the field and on the farm: principles and applications in research practices. Yale University Press New Haven, 13–34.
Jikomes N, Zoorob M (2018) The cannabinoid content of legal cannabis in Washington State varies systematically across testing facilities and popular consumer products. Scientific Reports 8 (1): 4519. https://doi.org/10.1038/s41598-018-22755-2
Knight G, Hansen S, Conner M, Poulsen H, McGovern C, Stacy J (2010) The result of an experimental indoor hyroponic Cannabis growing study using the ‘screen of green’ (ScOG) method: Yield, tetrahyrocannabinol (THC) and DNA analysis. Forensic Science International 202 (1–3): 36–44. https://doi.org/10.1016/j.forsciint.2010.04.022
Kottek M, Grieser J, Beck C, Rudolf B, Rubel F (2006) World map of Köppen-Geiger climate classification updated. Meteorologische Zeitscrift 15 (3): 259–263. https://doi.org/10.1127/0941-2948/2006/0130
Kudo Y, Kobayashi M, Momohara A, Noshiro S, Nakamura T et al. (2009) Radiocarbon dating of fossil hemp fruits in the earliest Jomon period. Shokuseishi kenkyū (Japanese Journal of Historical Botany) 17 (1): 27–32.
Lamarck JB (1785) Encyclopédie Méthodique, Botanique, Tome premier, part 2, Panckoucke, Paris, 345–752.
Lamb S (2010) In: Marcou D (Ed.) The smuggler’s ghost. Sirena Press, Madeira Beach, Florida.
Laverty KU, Stout JM, Sullivan MJ, Shah H, Gill N, Holbrook L, Deikus G, Sebra R, Hughes TR, Page JE, van Bakel H (2019) A physical and genetic map of Cannabis sativa identifies extensive rearrangements at the THC/CBD acid synthase loci. Genomic Research 29 (1): 146–156. https://doi.org/10.1101/gr.242594.118
Linnaeus C (1753) Species Plantarum, Tomus II. Laurentii Salvii, Holmiae [Stockholm].
Lynch RC, Vergara D, Tittes S, White K, Schwartz CJ, Gibbs MJ, Ruthenburg TC, deCesare K, Land DP, Kane NC (2016) Genomic and chemical diversity in Cannabis . Critical Reviews in Plant Sciences 35 (5–6): 349–363. https://doi.org/10.1080/07352689.2016.1265363
Mandolino G, Carboni A, Bagatta M, Cruicitti P, Ranalli P (2002) Variabilità e mappatura di marcatori molecolari RAPID in Cannabis sativa L. Agroindustria 1 (1): 3–7.
McPartland JM (1992) C.H. Persoon: A phanerogamic footnote. Mycotaxon 45 : 257–258. https://doi.org/10.1016/0007-1226(92)90099-J
McPartland JM (2017) “ Cannabis sativa and Cannabis indica versus “ Sativa ” and “ Indica ”. In: Chandra S, Lata H, ElSohly MA (Eds) Cannabis sativa : Botany and Biotechnology. Springer International Publishing, Cham, Switzerland , 101–121. https://doi.org/10.1007/978-3-319-54564-6_4
McPartland JM (2018) Cannabis systematics at the levels of family, genus, and species. Cannabis and Cannabinoid Research 3 (1): 203–212. https://doi.org/10.1089/can.2018.0039
McPartland JM, Hegman W (2018) Cannabis utilization and diffusion patterns in prehistoric Europe: A critical analysis of archaeological evidence. Vegetation History and Archaeobotany 27 (4): 627–634. https://doi.org/10.1007/s00334-017-0646-7
McPartland JM, Clarke RC, Watson DP (2000) Hemp diseases and pests – management and biological control. CABI Publishing, Wallingford, UK. https://doi.org/10.1079/9780851994543.0000
McPartland JM, Hegman W, Long TW (2019) Cannabis in Asia: Its center of origin and early cultivation, based on a synthesis of subfossil pollen and archaeobotanical studies. Vegetation History and Archaeobotany 28 (6): 691–702. https://doi.org/10.1007/s00334-019-00731-8
Mudge EM, Murch SJ, Brown PN (2018) Chemometric analysis of cannabinoids: Chemotaxonomy and domestication syndrome. Scientific Reports 8 (1): 13090. https://doi.org/10.1038/s41598-018-31120-2
Onofri C, de Meijer EPM, Mandolino G (2015) Sequence heterogeneity of cannabidiolic- and tetrahydrocannabinolic acid-synthase in Cannabis sativa L. and its relationship with chemical phenotype. Phytochemistry 116 : 57–68. https://doi.org/10.1016/j.phytochem.2015.03.006
Peterson C (2009) Prohibition and resistance: a socio-political exploration of the changing dynamics of the southern African cannabis trade, c. 1850-the present. Masters thesis, Rhodes University, Grahamstown, South Africa.
Piluzza G, Delogu G, Cabras A, Marceddu S, Bullitta S (2013) Differentiation between fiber and drug types of hemp ( Cannabis sativa L.) from a collection of wild and domesticated accessions. Genetic Resources and Crop Evolution 60 (8): 2331–2342. https://doi.org/10.1007/s10722-013-0001-5
Potter D (2009) The propagation, characterisation and optimisation of Cannabis sativa L. as a phytopharmaceutical. Doctoral thesis, King’s College, London.
Qu Y, Waley A (1955) The Nine Songs: a study of shamanism in ancient China. Allen and Uwin, London.
Ren M, Tang ZH, Wu XH, Spengler R, Jiang HG et al. (2019) The origins of cannabis smoking: chemical residue evidence from the first millennium BCE in the Pamirs. Science Advances 5: eaaw1391. https://doi.org/10.1126/sciadv.aaw1391
Sawler J, Stout JM, Gardner KM, Hudson D, Vidmar J, Butler L, Page JE, Myles S (2015) The genetic structure of marijuana and hemp. PLoS One 10(8): e0133292. https://doi.org/10.1371/journal.pone.0133292
Schultes RE, Klein WM, Plowman T, Lockwood TE (1974) Cannabis : An example of taxonomic neglect. Harvard University Botanical Museum Leaflets 23 : 337–367.
Schwabe AL, McGlaughlin ME (2018) Genetic tools weed out misconceptions of strain reliability in Cannabis sativa : implications for a budding industry. BioRxiv Preprint. https://doi.org/10.1101/332320
Seedfinder (2019) Seedfinder online database. https://en.seedfinder.eu/
Seidel U (1989) Studien zum Vokabular der Landwirtschaft im Syrischen II. Altorientalische Forschungen 16 (1–2): 89–139. https://doi.org/10.1524/aofo.1989.16.12.89
Sharma S (1988) Drive against charas launched. Tribune India 14 October 1988.
Small E (1972) Interfertility and chromosomal uniformity in Cannabis . Canadian Journal of Botany 50 (9): 1947–1949. https://doi.org/10.1139/b72-248
Small E (1975) Morphological variation of achenes of Cannabis . Canadian Journal of Botany 53 (10): 978–987. https://doi.org/10.1139/b75-117
Small E (2007) Cannabis as a source of medicinals, nutraceuticals, and functional foods. In: Acharya SN, Thomas JE (Eds) Advances in Medicinal Plant Research. Research Signpost, Trivandrum, Kerala, India , 1–39.
Small E (2015) Evolution and classification of Cannabis sativa (marijuana, hemp) in relation to human utilization. Botanical Review 81 (3): 189–294. https://doi.org/10.1007/s12229-015-9157-3
Small E (2017) Cannabis : a complete guide. CRC Press, Boca Raton, FL.
Small E (2018) Dwarf germplasm: The key to giant Cannabis hempseed and cannabinoid crops. Genetic Resources and Crop Evolution 65 (4): 1071–1107. https://doi.org/10.1007/s10722-017-0597-y
Small E, Beckstead HD (1973) Common cannabinoid phenotypes in 350 stocks of Cannabis . Lloydia 36 : 144–16.
Small E, Cronquist A (1976) A practical and natural taxonomy for Cannabis . Taxon 25 (4): 405–435. https://doi.org/10.2307/1220524
Small E, Jui P, Lefkovitch LP (1976) A numerical taxonomic analysis of Cannabis with special reference to species delimitation. Systematic Botany 1 (1): 67–84. https://doi.org/10.2307/2418840
Spitzer-Rimon B, Duchin S, Bernstein N, Kamenetsky R (2019) Architecture and florogenesis in female Cannabis sativa plants. Frontiers of Plant Science 10: 350. https://doi.org/10.3389/fpls.2019.00350
Takhtajan AL (1986) In: Crovello TJ transl, Cronquist A (Eds) Floristic regions of the world. University of California Press, Berkeley.
Turland NJ (2018) International Code of Nomenclature for Algae , Fungi , and Plants (Shenzhen Code). Koeltz Scientific Books, Königstein, Germany.
Turner CE, Hadley K, Fetterman PS (1973) Constituents of Cannabis sativa L. VI. Propyl homologs in samples of known geographical origin. Journal of Pharmaceutical Sciences 62 (10): 1739–1741. https://doi.org/10.1002/jps.2600621045
Turner CE, Cheng PC, Lewis GS, Russell MH, Sharma GK (1979) Constituents of Cannabis sativa . XV: Botanical and chemical profile of Indian variants. Planta Medica 37 (11): 217–225. https://doi.org/10.1055/s-0028-1097331
Turner CE, ElSohly MA, Boeren EG (1980) Constituents of Cannabis sativa L. XVII. A review of the natural constituents. Journal of Natural Products 43 (2): 169–234. https://doi.org/10.1021/np50008a001
Van Bakel H, Stout JM, Cote AG, Tallon CM, Sharpe AG, Hughes TR, Page JE (2011) The draft genome and transcriptome of Cannabis sativa . Genome Biology 12(10): R102. https://doi.org/10.1186/gb-2011-12-10-r102
Vavilov NI (1922) Полевые культуры Юго-Востока. Труды по прикладной ботанике, генетике и селекции 13 (Suppl. 23): 147–148.
Vavilov NI (1926) Происхождение культурной конопли и воэникновение культуры группы «первичных» растений. Труды по прикладной ботанике, генетике и селекции 16 (2): 107–121.
Vavilov NI (1935) “The phyto-geographical basis for plant breeding”. Теоретические основы селекции растений, Том 1. Sel’chozgiz, Moscow. Reprinted in Vavilov (1992), 316–366.
Vavilov NI (1992) In: Dorofeyev VF (Ed.) Origin and Geography of Cultivated Plants. Cambridge University Press, Cambridge UK.
Vavilov NI, Bukinich DD (1929) Konopli. Труды по прикладной ботанике, генетике и селекции 33 (Suppl.): 380–382.
Watson DP (1985) Cultivator’s choice catalog #4. Self-published, Amsterdam, Holland.
Weiblen GD, Wenger JP, Craft KJ, ElSohly MA, Mehmedic Z, Treiber EL, Marks MD (2015) Gene duplication and divergence affecting drug content in Cannabis sativa . The New Phytologist 208 (4): 1241–1250. https://doi.org/10.1111/nph.13562
White F, Léonard J (1991) Phytogeographical links between Africa and southwest Asia. Flora et Vegetatio Mundi 9 : 229–246.
Wiegand KM (1935) A taxonomist’s experience with hybrids in the wild. Science 81 (2094): 161–166. https://doi.org/10.1126/science.81.2094.161
Zinger NV (1898) Beiträge zur Kenntnis der weiblichen Blüthen und Inflorescenzen bei Cannabineen. Flora oder Allgemeine Botanische Zeitung 85 : 189–253.
Magyar fordítás és szerkesztési megjegyzés
Magyar fordítás és ábraszöveg-adaptáció. A fordítás az eredeti szerzők, a forrás, a DOI és a licenc feltüntetésével készült; a változtatás a magyar nyelvű fordítás és szerkesztés.
Kapcsolódó tanulmányok
Genomikai-taxonómiai áttekintés 2023 · Pángenomikai-evolúciós értelmezési modell 2025